| Hydrophilidae[1] | |||
| Latreille, 1802 | |||
| Okres istnienia: jura–dziś | |||
_female_(4035156238).jpg.webp) Kałużnica czarnozielona, samica | |||
| Systematyka | |||
| Domena | |||
|---|---|---|---|
| Królestwo | |||
| Typ | |||
| Gromada | |||
| Rząd | |||
| Podrząd | |||
| Infrarząd | |||
| Nadrodzina | |||
| Rodzina |
kałużnicowate | ||
| Synonimy | |||
| |||
Kałużnicowate (Hydrophilidae) – rodzina chrząszczy z podrzędu wielożernych i nadrodziny kałużnicokształtnych. Rozmieszczona kosmopolitycznie. Obejmuje około 2900 opisanych gatunków. Zamieszkują środowiska wodne lub środowiska lądowe o dużej zawartości wody, rzadko suchsze. Postacie dorosłe są detrytusożerne, roślinożerne lub wszystkożerne, wyjątkowo pyłkożerne. Larwy są drapieżnikami, zwykle polującymi z zasadzki. Postacie dorosłe i większość larw oddycha powietrzem atmosferycznym, tylko nieliczne larwy oddychają skrzelotchawkami lub powierzchnią ciała. Wodne imagines zwykle wykorzystują w celach oddechowych pęcherzyk powietrza utrzymywany plastronem. Owady dorosłe wykorzystują do komunikacji i odstraszania napastników strydulację. Z wyjątkiem jednego gatunku rozmnażają się wyłącznie płciowo. Przepoczwarczenie następuje niemal zawsze na lądzie. W zapisie kopalnym grupa w sposób pewny reprezentowana jest od jury późnej, natomiast z kredy wczesnej znane są już niektóre plemiona współczesne.
Morfologia
Owad dorosły

Chrząszcze o mniej lub bardziej zaokrąglonym w zarysie, przeciętnie lub silnie wysklepionym ciele długości od 0,9 do 46 mm, ale tylko w przypadku Hydrophilini przekraczające 10 mm[2]. Zarys ciała nie jest między przedpleczem a pokrywami przerwany lub przerwa ta jest słabo zaznaczona[3]. Ubarwienie mają żółtawe, brązowe lub czarne, tylko u nielicznych rodzajów z połyskiem metalicznym. Wierzch ciała zwykle jest gładki i nagi, co najwyżej miejscami z grupkami trichobotrii, ale niekiedy bywa owłosiony. Spód ciała zaopatrzony jest w plastron w postaci hydrofobowego owłosienia, czasem częściowo zredukowany, rzadko całkiem zanikły[2].
Głowa jest prognatyczna, o silnie poprzecznej wardze górnej, dużym i wypukłym nadustku, złożonych z drobnych omatidiów oczach i często dobrze zaznaczonym szwie epistomalnym[2][3]. Skronie są za oczami zwykle słabo zwężone i rzadko odgraniczone od nich listewkami[3]. Czułki zbudowane są z od siedmiu do dziewięciu członów, z których trzy ostatnie formują poprzedzoną kupulą owłosioną buławkę. Z wyjątkiem Omicrini i Megasternini nasady czułków zakryte są nadustkiem. Żuwaczki mają dobrze wykształcone mole i prosteki oraz jedno- lub dwuzębne wierzchołki; tylko u niektórych Omicrini żuwaczki są zredukowane. Szczęki mają oszczecinione żuwki, z których zewnętrzna jest spłaszczona, a wewnętrzna krótka i w dużej mierze błoniasta. Głaszczki szczękowe u niektórych grup są tak długie jak czułki lub jeszcze dłuższe, ale u Cyclominae i gomolatek są znacznie od czułków krótsze. Warga dolna typowo ma poprzeczną bródkę, dwupłatowy języczek i trójczłonowe głaszczki wargowe o zdrobniałym członie pierwszym[2]. Szwy gularne są prawie równoległe i w przedniej połowie bliskie sobie[3]. Szyja ma dobrze wykształcone skleryty szyjne[2].
Przedplecze jest wyraźnie szersze niż dłuższe, najszersze pośrodku lub w tyle, niemal zawsze mniej więcej tak szerokie jak podstawy pokryw, o zaokrąglonych kątach przednich i zaokrąglonych lub prostych kątach tylnych[2], niemal zawsze równomiernie wysklepione i zazwyczaj równomiernie punktowane[3]. Boki ma zakrzywione, o kompletnych i niezmodyfikowanych listewkach krawędziowych. Tylny brzeg jest prosty do lekko falistego i zaopatrzony od spodu w listewkę. Tarczka jest z tyłu zaostrzona[2], zawsze dobrze widoczna[3]. Pokrywy mają dobrze zaznaczone barki, zwykle po od 9 do 11 rzędów punktów, czasem jednak rzędy są nieregularne lub punktowanie jest bezładne; u części grup występują ponadto rządki przytarczkowe. Wierzchołki pokryw z wyjątkiem rodzaju gomolatka są wspólnie zaokrąglone. Epipleury zwykle są zredukowane w tyle, ale bywają też na całej długości bardzo szerokie[2]. Skrzydła tylnej pary są zwykle najszersze przed pętlą mediokubitalną (rzadziej dalej), o ostrodze kubitalnej wyrastającej ze szczytu wspomnianej pętli, zwykle o żyłce medialnej widocznej także na pewnym odcinku przed żyłką poprzeczną radialno-medialną, z wyjątkiem Megasternini o wyraźnej komórce bazalnej[3].
Przedtułów ma hypomery podzielone na nagie części boczne i owłosione części dośrodkowe, niekiedy odseparowane listewkami. Przedpiersie ma słabo wykształcony wyrostek międzybiodrowy. Panewki bioder przednich są od zewnątrz szeroko otwarte, od wewnątrz zwykle styczne, rzadko wąsko odseparowane. Zazwyczaj kompletne szwy anapleuralne odgradzają śródpiersie od mezanepisternitów. Najczęściej pośrodku śródpiersia znajduje się żeberko lub wyniesiona płytka. Panewki środkowych bioder zwykle są wąsko odseparowane. Poprzeczne zapiersie zwykle jest płaskie do lekko sklepionego, u niektórych gomolatek jednak bardziej pośrodku sklepione[2].

Biodra przedniej pary są stożkowate i wydatne, środkowej poprzeczne i ukośne (tylko u Amphiops niemal kuliste[3]) z odsłoniętymi krętarzykami, a tylnej silnie poprzeczne i niemal stykające się. Krętarze łączą się z udami po skosie[2]. Uda zwykle mają rowki do chowania goleni[3]. Kształt goleni bywa od walcowatego po silnie spłaszczony; nie występuje piłkowanie. U gatunków wodnych golenie drugiej i trzeciej pary (rzadko tylko drugiej) czasem zaopatrzone są w włoski pływne. Stopy są pięcioczłonowe, rzadko czteroczłonowe w przypadku ostatniej ich pary, zazwyczaj o dwuszczecinkowym (rzadko wieloszczecinkowym) empodium[2][3].
Na spodzie odwłoka widocznych jest zwykle pięć wolnych sternitów (od trzeciego do siódmego), rzadko sześć lub cztery[2][3]. Tylny brzeg siódmego sternitu jest zaokrąglony lub na szczycie wcięty, rzadko z ząbkiem[3]. Funkcjonalne przetchlinki występują na segmentach od pierwszego do szóstego[2]. Pod pokrywami na trzecim laterosternicie obecne są elementy aparatu strydulacyjnego, zwykle w postaci szeregów guzków, rzadziej pozlewanych w pełne listewki. U samca edeagus typowo składa się z fallobazy, pary niezmodyfikowanych paramer i płata środkowego (prącia). Genitalia samic mają słabo zesklerotyzowane pokładełko o krótszych od koksytów paraproktach[2].
Stadia rozwojowe
Jajo
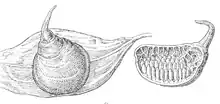
Jaja umieszczane są zwykle w kapsułach jajowych o rozmaitych, charakterystycznych dla rodzajów lub gatunków kształtach. Kapsułki utworzone są z jedwabistej przędzy i zwykle dwuwarstwowe, jednowarstwowe tylko u Anacaena. Często kapsułka zaopatrzona jest w maszt, zwykle taśmowaty, którego funkcja pozostaje niepewna[3].
Larwa

Larwy mają ciało pociągłe do nieco pośrodku rozszerzonego, walcowate lub lekko spłaszczone[2].
Głowa jest hiperprognatyczna[2][3], rzadziej prognatyczna[3], dobrze zesklerotyzowana[2][3], nieco spłaszczona grzbietobrzusznie, prawie owalna do prawie czworokątnej w zarysie[3], o V-kształtnych ramionach szwu epikranialnego i co najwyżej krótkim jego trzonie, pozbawiona szwu epistomalnego. Warga górna zlana jest z nadustkiem w klipeolabrum o przedniej krawędzi zaopatrzonej w nasale (rzadko o nasale zredukowanym lub całkiem zanikłym[3]) i adnasalia[2]. Osadzone po grzbietowej stronie głowy[3] czułki są trój-, rzadko czteroczłonowe (wskutek podziału członu drugiego), o członie drugim zaopatrzonym w pojedynczy, boczny wyrostek zmysłowy[2][3]. Żuwaczki są sierpowate, zwykle o mającym od jednego do trzech ząbków retynakulum, pozbawione penicillusa i moli. Bruzda szczękowa jest płytka[3]. Sześcioczłonowe szczęki mają kotwiczkę w formie pojedynczego sklerytu, długi i walcowaty pieniek, członopodobny dźwigacz, małą żuwkę zewnętrzną i trójczłonowy głaszczek każda; brak jest żuwki wewnętrznej[2]. Warga dolna ma prawie trójkątny do prawie pięciokątnego podbródek przyrośnięty do puszki głowowej, prawie czworokątną bródkę, prawie prostokątny przedbródek i dwuczłonowe głaszczki, pozbawiona jest zaś języczka[2][3]. Szwy gularne są prawie zlane, a skleryt guli zredukowany. Most tentorialny jest U-kształtny lub zanikły, a tylne bruzdy tentorialne niemal styczne[3].
Przedtułów ma silnie zesklerotyzowane przedplecze, a na spodzie podzieloną lub pełną płytkę przedpiersia. Śródtułów i zatułów są zwykle krótsze od przedtułowia i słabiej sklerotyzowane, o tergitach mniejszych, prawie owalnych do prawie trójkątnych. Odnóża bywają wykształcone rozmaicie, od pięcioczłonowych przez trójczłonowe po całkiem zanikłe[2][3]. Biodra przedniej pary są rozstawione szeroko, pozostałych zaś par wąsko[2].
Odwłok zbudowany jest z dziesięciu segmentów, z których osiem pierwszych jest słabo zesklerotyzowanych, zaopatrzonych w dwa lub więcej drobnych sklerytów. Segmenty od ósmego do dziesiątego zwykle formują przedsionek przetchlinkowy. Dziewiąty segment, z wyjątkiem trzech rodzajów, ma parę małych, jednoczłonowych urogomfów, a dziesiąty jest zredukowany i umieszczony brzusznie[2][3]. U leniwczyków występują po bokach odwłoka skrzelotchawki[3].
Poczwarka

Poczwarki są białawe, rzadko ciemne, a u Derallus zielonkawe. Na wierzchu głowy i wszystkich segmentów tułowia oraz na grzbiecie i bokach segmentów odwłoka od pierwszego do siódmego lub ósmego występują wyrostki oskórkowe (styliki), z wyjątkiem rodzaju Amphiops opatrzone na szczycie szczecinką. Funkcjonalne przetchlinki leżą na segmentach odwłoka od drugiego do szóstego. Segment dziewiąty dysponuje parą przysadek odwłokowych[2][3].
Biologia i ekologia
Siedlisko
Chrząszcze pierwotnie wodne, ale liczne linie ewolucyjne zaadaptowały się do środowisk lądowych, a niektóre z nich wróciły do wody. Gatunki wodne zamieszkują rozmaite wody płynące i stojące, także o okresowym charakterze, chętnie o bogatej szacie roślinnej; niektóre wyspecjalizowane są w fitotelmach, mokrych skałach (gatunki higropetryczne) czy wysiękach. Związane są z wodami słodkimi[2][3], rzadko ze słonawymi[4]. Część gatunków jest półwodnych lub związanych z pobrzeżami wód. Te zasiedlające rozkładające się na wybrzeżach morskich wodorosty uodporniły się na okresowe zanurzanie w słonej wodzie. Formy lądowe zwykle zamieszkują siedliska wilgotne, z dużą zawartością wody, w tym wilgotną ściółkę, butwiejące drewno, odchody ssaków, a rzadziej ptaków, padlinę, wilgotne płaty mchów, gnijące owoce i inne szczątki roślinne, owocniki grzybów i śluzowców czy wycieki soków z drzew. Nieliczne kałużnicowate znajdywane są w siedliskach suchszych, np. w mrowiskach, termitierach i na kwiatach. Formy lądowe dominują w podrodzinie gomolatek, a w pozostałych podrodzinach większość stanowią formy wodne[2][3].
W Europie bentosowe kałużnicowate uznane zostały na potrzeby punktacji w systemie BMWP za charakterystyczne dla wód o przeciętnych parametrach jakościowych[5].
Interakcje międzygatunkowe
Pokarm



Postacie dorosłe w większości przypadków żerują na rozkładającej się materii organicznej, rzadziej na żywych glonach czy roślinach wyższych. Zwykle dla udanego rozrodu nawet gatunki niedrapieżne wymagają uzupełnienia diety białkiem zwierzęcym, stąd sporadycznie żerują na padlinie, a nawet żywych zwierzętach. Największe osobniki są w stanie polować na ślimaki, kijanki czy małe ryby. Istotnym wyjątkiem jest Rygmondus, którego owady dorosłe żerują na pyłku (melitofagia)[2][3].
Larwy są drapieżnikami, zwykle polującymi z zasadzki. Ich ofiarami padają owady, skorupiaki, pierścienice, a w przypadku większych osobników także ślimaki, kijanki i małe ryby; wyjątkiem są tylko niektóre leniwczyki żerujące na glonach. W wyborze ofiary są oportunistami i częsty jest wśród nich kanibalizm; w warunkach naturalnych przeciwdziała mu częściowo krótka faza dyspersyjna po wylęgu, nieobecna jednak u koprofili. Wykorzystują trawienie pozaustrojowe – soki trawienne wylewane są przez lejkowaty aparat gębowy do jamy przedgębowej i po wstępnym strawieniu ofiary powstała pulpa jest zasysana do jelit, a twarde elementy odrzucane. W przypadku gatunków wodnych znane są dwie strategie radzenia sobie z problemem rozcieńczania soków przez środowisko wodne. Najpopularniejszą jest wyniesienie ofiary ponad taflę wody, utrzymując ją i manipulując nią za pomocą czułków, narządów gębowych, a czasem też przedniej pary odnóży – to właśnie do takiego sposobu żerowania stanowi przystosowanie hiperprognatyczna głowa. Drugą, stosowaną przez larwy bentosowe, strategią jest dociskanie ofiary do odwłoka, przebicie jej oskórka dwiema żuwaczkami lub tylko lewą z nich oraz wstrzyknięcie do jej wnętrza soków trawiennych i późniejsze wypicie rozpuszczonych wnętrzności przez bruzdę w żuwaczce. Larwy z rodzaju kałużnica potrafią ponadto dobierać się do ślimaków – dociskają muszlę uniesioną głową do grzbietowej strony odwłoka i rozgniatają ją żuwaczkami[2][3]. Muszla może też służyć do zabezpieczenia soków trawiennych przed rozcieńczeniem, dzięki czemu ślimaki mogą być zjadane bez wynurzania[2].
Drapieżniki i pasożyty
Na kałużnicowate polują ryby, ptaki, żółwie oraz bezkręgowce, zwłaszcza pluskwiaki wodne, wodne chrząszcze drapieżne i larwy ważek. Niektóre gatunki jadane są także przez ludzi[3].
Do pasożytów kałużnicowatych należą grzyby (w tym owadorostowce), nicienie, gregaryny i roztocza. Parazytoidami są natomiast niektóre owadziarki, muchówki i chrząszcze z podrodziny strzelowatych[3].
Forezja i mutualizm
Foretycznie na ciele kałużnicowatych występują kolonie pierwotniaków i niektóre roztocza. Wyrostki na ciele larw niektórych gatunków porastane są przez glony, co usprawnia ich kamuflaż[3].
Ruch
Osobniki dorosłe są zwykle dobrymi lotnikami, ale po powierzchniach przemieszczają się mniej sprawnie. Większość gatunków wodnych łazi po podłożu lub zanurzonych roślinach i niezbyt dobrze pływa. Lepszymi pływakami są tylko niektórzy przedstawiciele Hydrophilinae, ale nawet oni pływają słabiej od pływakowatych czy krętakowatych. Chrząszcze pływają machając na przemian prawymi i lewymi odnóżami środkowej i tylnej pary[3].
Wśród larw gatunków wodnych również dominują te łażące po dnie lub roślinach. Dobrze pływają jedynie larwy Hydrophilini, które mają odnóża tylnej i środkowej pary zaopatrzone w długie włoski pływne na udach i stopogoleniach. U części gatunków lądowych larwy mają odnóża uwstecznione lub całkiem zanikłe, co stanowi adaptację do życia w gnijących szczątkach roślinnych czy odchodach. Niektóre larwy jako mechanizm obronny stosują szybkie, wykręcające ciało ruchy[3].
Oddychanie
Postacie dorosłe oddychają powietrzem atmosferycznym. Gatunki wodne na spodzie ciała utrzymują dzięki hydrofobowym włoskom (plastronowi) pęcherzyk powietrza, który uzupełniają przy użyciu również wyposażonych w hydrofobowe owłosienie czułków. Wyjątkiem są tylko niektórzy przedstawiciele rodzaju Hydrophilus o plastronie odwłoka zredukowanym. Plastron częściowo zachowany jest również u większość gatunków lądowych[2][3].
Larwy większości gatunków również oddychają powietrzem atmosferycznym. Dominują larwy metapneustyczne – o dużych, funkcjonalnych przetchlinkach umieszczonych na ósmym segmencie odwłoka i zamkniętych w przedsionku utworzonym przez z modyfikacji segmentów od ósmego do dziesiątego. Przedsionek ten otwiera się, kiedy larwa wystawia czubek odwłoka ponad taflę by czerpać powietrze i zamyka, gdy larwa się zanurza. Struktura ta zachowana jest u większości form lądowych, co świadczy o pierwotnie wodnym środowisku życia kałużnicowatych. Wodne larwy części gatunków wciągają ponadto powietrze atmosferyczne do jelit, gdzie tworzy ono pęcherzyk poprawiający wyporność. Rozwiązania tego nie stosują larwy dobrze pływające oraz bentosowe. U dwóch wodnych linii ewolucyjnych pojawiły się larwy apneustyczne, oddychające pod wodą za pomocą umieszczonych po bokach odwłoka skrzelotchawek o bardzo cienkim oskórku[2][3]. U rodzajów Hemiosus i Hybogralius wymiana gazowa następuje poprzez powierzchnię ciała – brak skrzelotchawek, a przedsionek jest niefunkcjonalny u pierwszego i zanikły u drugiego[2][3].
Strydulacja
Postacie dorosłe potrafią wydawać dźwięki poprzez pocieranie plectrum pokryw o guzki lub listewki na trzecim laterosternicie odwłoka[2]. Występująca u postaci dorosłych strydulacja służy zarówno jako mechanizm obronny, jak i do komunikacji między osobnikami, przy czym w celach tych wykorzystywane są różne dźwięki. Ponadto w obrębie dźwięków komunikacyjnych przynajmniej u części gatunków istnieje rozróżnienie na dźwięk wabiący samicę, dźwięki używane przez samca i samicę w trakcie zalotów, jak wydawany przez niezainteresowaną samicę dźwięk odmawiający zalotów[2][3].
Rozród i rozwój
.jpg.webp)
.jpg.webp)
_Figure_18.jpg.webp)
_(2977745747).jpg.webp)
_(2978610110).jpg.webp)
.jpg.webp)
_Figure_28.jpg.webp)
_(3052939128).jpg.webp)
_Figure_30.jpg.webp)
Niemal wszystkie kałużnicowate rozmnażają się płciowo. Tylko u Anacaena lutescens występuje partenogeneza, połączona z triploidią i polimorfizmem[3].
Samica po zaplemnieniu składa jaja do struktur utkanych z przędzy, pojedynczo lub po kilka sztuk. Struktury te zwykle mają formę kapsuły (torebki) z masztem służącym prawdopodobnie wymianie gazowej. Jedno zaplemnienie wystarcza do produkcji większej ilości torebek, aczkolwiek samice często kopulują wielokrotnie podczas jednego okresu rozrodczego. Gatunki wodne przyklejają torebki do roślin i różnych zanurzonych powierzchni, rzadziej torebki jajowe unoszą się na powierzchni wody z masztem sterczącym ku górze, a u niektórych rodzajów torebka noszona jest przez samicę do wylęgu larw. U nielicznych taksonów jaja składane są do luźnej, nieukształtowanej w kapsułę przędzy[2][3].
Larwy klują się zwykle po 4 do 8 dniach od złożenia jaj, ale u koprofilnych gomolatek już po 2–3 dniach, a u Tropisternus dopiero po dwóch tygodniach. Rozwój larwalny zajmuje typowo od 40 do 60 (rzadko 90) dni i obejmuje trzy stadia. U koprofilnych gomolatek rozwój larwalny trwa jednak od 7 do 20 dni, a stadia mogą być tylko dwa. Wyrośnięta larwa wchodzi w krótką (zwykle trwającą jedną lub dwie doby) fazę przedpoczwarkową, w której nie pobiera pokarmu i zajmuje się tylko znalezieniem miejsca do przepoczwarczenia[2][3].
Przepoczwarczenie następuje niemal zawsze na lądzie, w komorze wykopanej w wilgotnym podłożu lub skonstruowanej ze szczątków roślinnych; niektórzy przedstawiciele rodzaju Enochrus przepoczwarczają się w wodzie, w komorze z glonów, a niektórzy przedstawiciele rodzaju Cymbiota wśród mchów; tylko u rodzajów Derallus i Amphiops poczwarki są wolne i przyklejone do roślin wodnych. Stadium poczwarki zwykle zajmuje od 5 do 7 dni, ale u dwóch wspomnianych rodzajów skrócone jest do około dwóch dób. Zwykle opuszczające poczwarki osobniki dorosłe, często jeszcze w fazie teneralnej, rozlatują się w poszukiwaniu nowych siedlisk (dyspersja)[2][3].
Rozprzestrzenienie
Rodzina kosmopolityczna, znana ze wszystkich krain zoogeograficznych[2][3][4]; jej przedstawicieli brak jedynie na Antarktydzie. Kilkanaście rodzajów ma również zasięgi kosmopolityczne, większość ograniczona jest jednak do jednej lub dwóch krain. Kosmopolityczna jest większość podrodzin; wyjątkami są Enorchinae i Cylominae – tych pierwszych brak w krainie australijskiej, a tych drugich w Holarktyce[3]. Spośród plemion niekosmopolityczne zasięgi mają Amphiopini, których brak w Amerykach, oraz Protosternini, ograniczone do krainy orientalnej i północy krainy australijskiej[3][4]. W Polsce stwierdzono 77 gatunków[6] (zobacz: kałużnicowate Polski).
Taksonomia i ewolucja
Koncepcje historyczne
Takson ten wprowadził w 1802 roku Pierre André Latreille pod nazwą Hydrophilii, osobno wprowadzając takson Sphaeridiota dla gomolatek[7][8]. Na przestrzeni wieków rodzina kałużnicowatych była różnie definiowana i dzielona[4]. W 1815 roku William Elford Leach podzielił Hydrophilii na obejmujące rodzaje Hydrophilus, Hydrous, Hydrobius, Limnebius i Spercheus Hydrophilida oraz obejmujące rodzaje Helophorus, Ochtebius i Hydraena Helopherida[9]. W 1817 roku Leach ujął rodziny Hydrophilidea, Helophoridea i Sphaerididea w obrębie wyższego taksonu Palpicornia[10]. W 1837 roku Wilhelm Ferdinand Erichson do tych trzech taksonów dodał jeszcze Spercheini i określił je jako Hydrophilen[11].
Pierwszą szczegółową klasyfikację kałużnicowatych stworzył w 1844 roku Étienne Mulsant[12][4] – w obrębie kałużnic wyróżnił Hélophoraries oraz Hydrophilines z pięcioma grupami: Limnébiaires, Bérosaires, Cyllidiaires, Hydrobiaires i Hydrophilaires, osobno jednak wyróżniając gomolatki jako Sphéridiens z grupami Sphéridiaires i Mégasternaires[12][13]. Przez resztę XIX wieku i początek wieku XX wieku dokładane były do tego systemu kolejne taksony ponadrodzajowe[13].
Kolejną kompleksową, mającą odzwierciedlać filogenezę, systematykę kałużnic stworzył w pracach z 1916 i 1919 roku Armand d'Orchymont. W obrębie kałużnicokształtnych (Palpicornia) wyróżnił dwie kategorie, z których druga obejmowała Helophorinae, Epimetopinae, Hydrochinae, Sphaeridiinae i Hydrophilinae. Sphaeridiinae dzielił na plemiona Rygmodini, Sphaeridiini i Cercyonini, a Hydrophilinae na plemiona Hydrobiini, Hydrophilini, Amphiopini i Berosini z podplemionami Hydrobiae i Helocharae[14][15][13]. Podział taki z niewielkimi modyfikacjami przyjmowany był przez większą część XX wieku[13]. W pracach z wieku XIX i XX widać ogólnie rozbicie współcześnie rozumianych kałużnicowatych pomiędzy głównie lądowe „Sphaeriidines” i głównie wodne „Hydrophilines”[4].
Przełom XX i XXI wieku
W 1991 roku Michael Hansen opublikował wyniki pierwszej, opartej głównie na morfologii postaci dorosłych, całościowej analizy filogenetycznej nadrodziny kałużnicokształtnych i opartą na niej klasyfikację[4][13]. Podzielił on Hydrophiloidea na rodziny oguzkowatych, Epimetopidae, ziemioryjkowatych, wodopływkowatych, Spercheidae oraz kałużnicowatych. W obrębie tej ostatniej wyróżniał podrodziny Horelophinae, Hydrophilinae z plemionami Sperchopsini, Berosini, Chaetarthriini, Anacaeniini, Oocyclini, Hydrophilini (z trzema podplemionami) oraz podrodzinę Sphaeridiinae z plemionami Rygmodini, Tormissini, Andotypini, Borborophorini, Coelostomatini, Protosternini, Omicrini, Megasternini i Sphaeridiini[13]. W 1997 roku Hansen dodał jeszcze do kałużnicowatych podrodzinę Horelophopsinae[16]. Klasyfikacja taka była na początku XXI wieku stosowana przez większość specjalistów od chrząszczy wodnych i została utrwalona w licznych katalogach, także fauny światowej[4]. Niektóre źródła ogólne stosowały jednak system z szerszej zdefiniowanymi kałużnicowatymi[4][8]; np. w klasyfikacji chrząszczy Patrice’a Boucharda i innych z 2011 roku do kałużnicowatych zaliczono podrodziny Helophorinae, Epimetopinae, Georissinae, Hydrochinae, Spercheinae, Horelophinae, Horelophopsinae, Hydrophilinae z plemionami Anacaenini, Berosini, Chaetarthriini, Hydrophilini, Laccobiini i Sperchopsini oraz Sphaeridiinae z takimi samymi plemionami co w systemie Hansena[8].
Koncepcja współczesna
Po pracy Hansena ukazywały się kolejne filogenetyczne analizy morfologiczne i molekularne kałużnicokształnych (kilkanaście do 2013 roku), a niektóre cechy budowy podane przez Hansena zreinterpretowano[4]. Pierwszą analizą morfologiczno-molekularną tej nadrodziny była ta opublikowana w 2009 roku przez Detlefa Bernharda i współpracowników[17]. Znaczącej rewizji podziału kałużnicowatych dokonali w 2013 roku Andrew Edward Short i Martin Fikáček na podstawie obszernej analizy molekularnej. Wyróżnili w obrębie tej rodziny sześć podrodzin i 12 plemion[4]:
- Hydrophilinae Latreille, 1802
- Amphiopini Kuwert, 1890
- Berosini Mulsant, 1844
- Laccobiini Houlbert, 1922
- Hydrophilini Latreille, 1802
- Hydrobiusini Mulsant, 1844
- Chaetarthriinae Bedel, 1881
- Chaetharthriini Bedel, 1881
- Anacaenini Hansen, 1991
- Enochrinae Short et Fikáček, 2013
- Acidocerinae Zaitzev, 1908
- Cylominae Zaitzev, 1908
- Sphaeridiinae Latreille, 1802 – gomolatki
- Omicrini Smetana, 1975
- Coelostomatini Heyden, 1891
- Protosternini Hansen, 1991
- Sphaeridiini Latreille, 1802
- Megasternini Mulsant, 1844
Łącznie taksony te obejmują około 2900 opisanych gatunków[2].
Filogeneza
Monofiletyzm kałużnicowatych ma silne wsparcie w wynikach morfologicznych i molekularnych analiz filogenetycznych[2], np. Michaela Hansena z 1991 roku[13], Miguela Archangelskiego z 1998 roku[18], Rolfa G. Beutela i Albrechta Komarka z 2004 roku[19], Anji Korte i innych z 2004 roku[20], Detlefa Bernharda i innych z 2006 i 2009 roku[21][17], Branki B. Mađarić i innych z 2013 roku[22], Andrew E. Shorta i Martina Fikáčka z 2013 roku[4], Ladislava Bocáka i innych z 2014 roku[23] czy Duane D. McKenny i innych z 2015 roku[24]. Znane są synapomorfie grupy w budowie larw i owadów dorosłych[2]. Jako autapomorfie w morfologii imagines postuluje się pojedynczą, długą szczecinkę na ostatnim członie głaszczków wargowych, równomierną powierzchnię przedplecza, tworzenie przez boczny brzeg przedplecza ciągłej, wypukłej linii z brzegiem pokryw, pionowe epipleury, wygrodzone listewkami rowki do chowania goleni na udach, gęsto rozmieszczone kolce w najbardziej zewnętrznym szeregu na goleniach przedniej pary, długą szczelinę panewek bioder przednich czy mniej lub bardziej wyraźnie wyniesioną środkową część zapiersia[3]. Jako potencjalne autapomorfie w budowie larw wymienia się ścianę brzuszną puszki głowowej dłuższą od grzbietowej, skierowane ku górze narządy gębowe, ukośny otwór wielki, szczęki o zredukowanej bruździe umieszczone na tym samym poziomie co stawy żuwaczek, grzbietowo umieszczone czułki czy musculus tentoriopraementalis i musculus submentopraementalis wychodzące z V-kształtnej listwy lub apodemy na tylnym brzegu sklerytu podbrókowego[25].
Nie ma dotąd pewności co do tego, która rodzina jest z kałużnicowatymi spokrewniona najbliżej[2]. Na rzecz siostrzanej relacji kałużnicowatych i Spercheidae świadczą m.in. wyniki analiz Michaela Hansena z 1991 roku[13], Miguela Archangelskiego z 1998 roku[18], Rolfa G. Beutela i Albrechta Komarka z 2004 roku[19], Anji Korte i innych z 2004 roku[20], Branki B. Mađarić i innych z 2013 roku[22]. Klad utworzony przez Epimetopidae i Georissidae jako siostrzany dla kałużnicowatych pojawił się w wynikach Rolfa G. Beutela z 1994 roku[25]. Wyniki uzyskane przez Detlefa Bernharda i innych w 2006 roku umieszczają kałużnicowate jako siostrzane dla kladu obejmującego Epimetopidae, Hydrochidae, Georissidae i Helophoridae[21]. W publikacji Detlefa Bernharda i innych z 2009 roku Spercheidae jawiły się jako siostrzane dla kałużnicowatych przy użyciu metody bayesowskiej, natomiast przy użyciu metody największej parsymonii siostrzanym dla kałużnicowatych był klad obejmujący Georissidae, Epimetopidae, Hydrochidae i Helophoridae[17]. Epimetopidae jako grupę siostrzaną kałużnicowatych wskazują wyniki Andrew E. Shorta i Martina Fikáčka z 2013 roku[4]. W wynikach Ladislava Bocáka i innych z 2014 roku jako siostrzany dla kałużnicowatych rozpoznany został klad tworzony przez Helophoridae, Georissidae i Hydrochidae[23]. Wyniki analiz Duane D. McKenny i innych z 2015 roku wskazały Spercheidae jako siostrzane dla kałużnicowatych przy metodzie bayesowskiej, a klad obejmujący Georissidae i Helophoridae jako siostrzany dla kałużnicowatych przy metodzie największej wiarygodności[24].
Zapis kopalny

Analizy metodą zegara molekularnego wskazują na pojawienie się kałużnicokształtnych w triasie późnym[26][27][28]. Do nadrodziny tej przypisywano skamieniałości pojedynczych pokryw z tego okresu, jednak ich klasyfikacja jest niepewna[27]. Pewne skamieniałości kałużnicokształtnych pochodzą z jury i dotyczy to również kałużnicowatych[27][29][2]. Zapis kopalny tej rodziny jest dość obfity[29][2], tylko do 2010 roku opisano skamieniałości około 180 gatunków z 55 rodzajów, w tym 33 wymarłych. Z jury późnej znane są „Mesosperchus” schultzi i Protochares brevipalpis, a z kredy wczesnej należące do plemienia Hydrobiusini rodzaje Baissalarva i Cretoxenus, zaliczany do podrodziny Acidocerinae rodzaj Hydroyixia oraz należący do Acidocerinae lub Enochrinae rodzaj Alegorius. Przynależność kredowych skamieniałości do współczesnych plemion wskazuje na wysokie zróżnicowanie rodziny już w tym okresie[29].
Przypisy
- ↑ Hydrophilidae, [w:] Integrated Taxonomic Information System (ang.).
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 Martin Fikáček: 20. Hydrophilidae Leach, 1815. W: Adam Ślipiński, John F. Lawrence: Australian Beetles. Volume 2. Archostemata, Myxophaga, Adephaga, Polyphaga (part). CSIRO Publishing, 2019, s. 271-350. ISBN 978-0-643-09730-8.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 Miguel Archangelsky, Rolf Georg Beutel, Albrecht Komarek: 12.1 Hydrophilidae Latreille, 1802. W: Rolf G. Beutel, Richard A. B. Leschen: Arhtropoda: Insecta. Coleoptera, Beetles, Volume 1: Morphology and Systematics (Archostemata, Adephaga, Myxophaga, Polyphaga partim). 2nd edition. Berlin, Boston: Walter de Gruyter, 2016, s. 238-254, seria: Handbook of Zoology/Handbuch der Zoologie.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 Andrew Edward Z. Short, Martin Fikáček. Molecular phylogeny, evolution and classification of the Hydrophilidae (Coleoptera). „Systematic Entomology”. 38 (4), s. 723-752, 2013. The Royal Entomological Society. DOI: 10.1111/syen.12024.
- ↑ Barbara Bis, Artur Mikulec, Rajmund Jan Wiśniewski, Przewodnik do oceny stanu ekologicznego rzek na podstawie makrobezkręgowców bentosowych, Biblioteka Monitoringu Środowiska, Warszawa: Główny Inspektorat Ochrony Środowiska, 2013, s. 114, ISBN 978-83-61227-21-2 [dostęp 2024-03-13] [zarchiwizowane 2015-03-22].
- ↑ Marek Przewoźny: rodzina: Hydrophilidae Latreille, 1802 — kałużnicowate. [w:] Biodiversity Map [on-line]. [dostęp 2024-03-11].
- ↑ P.A. Latreille: Histoire naturelle, générale et particulière des crustacés et des insectes. Ouvrage faisant suite à l’histoire naturelle générale et particulière, composée par Leclerc de Buffon, et rédigée par C.S. Sonnini, membre de plusieurs sociétés savantes. Familles naturelles des genres. Tome troisième. Paris: F. Dufart, 1802.
- 1 2 3 Patrice Bouchard i inni, Family-group names in Coleoptera (Insecta), „ZooKeys”, 88, 2011, s. 1–972, DOI: 10.3897/zookeys.88.807.
- ↑ W.E. Leach, Entomology, [w:] D. Brewster (red.), Brewster’s Edinburgh Encyclopedia, t. IX, Edinburgh: W. Blackwood, J. Waugh, etc, 1815 (1), s. 57–172.
- ↑ William E. Leach: The zoological miscellany; being descriptions of new, or interesting animals. Illustrated with coloured figures, drawn from nature, by R.P. Nodder. Vol. III. London: E. Nodder & Sons, 1817.
- ↑ W.F. Erichson: Die Käfer der Mark Brandenburg. Erster Band. Erste Abtheilung. Berlin: F.H. Morin, 1837.
- 1 2 É. Mulsant, Palpicornes, [w:] Histoire naturelle des Coléoptères de France. [Tome 3], Paris: Maison, 1844.
- 1 2 3 4 5 6 7 8 Michael Hansen, The Hydrophiloid Beetles. Phylogeny, Classification and a Revision of the Genera (Coleoptera, Hydrophiloidea). Biologiske Skrifter No. 40, „Insect Systematics & Evolution”, 3, 23, Copenhagen: The Royal Danish Academy of Sciences and Letters & Munksgaard, 1991, DOI: 10.1163/187631292X00164, ISSN 1876-312X.
- ↑ A. d’Orchymont. Notes pour la classification et la phylogénie des Palpicornia. „Annales de Société entomologique de France”. 85, s. 91–106, 1916.
- ↑ A. d’Orchymont. Notes complémentaires pour la classification et la phylogénie des “Palpicornia”. „Revue Zoologique Africaine”. 6, s. 163–168, 1919.
- ↑ M. Hansen. A new subfamily for a remarkable new genus and species of Hydrophilidae from New Guinea (Coleoptera: Hydrophilidae). „Annales Zoologici”. 47, s. 107–110, 1997.
- 1 2 3 Detlef Bernhard, Rolf Beutel, Albrecht Komarek, Ignacio Ribera. Phylogenetic analysis of Hydrophiloidea (Coleoptera: Polyphaga) based on molecular data and morphological characters of adults and immature stages. „Insect Systematics and Evolution”. 40, s. 3-41, 2009. DOI: 10.1163/187631209X416741.
- 1 2 M. Archangelsky. Phylogeny of Hydrophiloidea (Coleoptera: Staphyliniformia) using characters from adult and preimaginal stages. „Systematic Entomology”. 23, s. 9–24, 1998.
- 1 2 R.G. Beutel, A. Komarek. Comparative study of thoracic structures of adults of Hydrophiloidea and Histeroidea with phylogenetic implications (Coleoptera, Polyphaga). „Organisms, Diversity and Evolution”. 4, s. 1–34, 2004.
- 1 2 A. Korte, I. Ribera, R. G. Beutel, D. Bernhard. Interrelationships of Staphyliniform groups inferred from 18S and 28S rDNA sequences, with special emphasis on Hydrophiloidea (Coleoptera, Staphyliniformia). „Journal of Zoological Systematics and Evolutionary Research”. 4, s. 281-288, 2004. Hindawi Limited. DOI: 10.1111/j.1439-0469.2004.00282.x.
- 1 2 D. Bernhard, C. Schmidt, A. Korte, G. Fritzsch, R.G. Beutel. From terrestrial to aquatic habitats and back again – molecular insights into the evolution and phylogeny of Hydrophiloidea (Coleoptera) using multigene analyses. „Zoologica Scripta”. 35, s. 597–606, 2006.
- 1 2 Branka Bruvo Mađarić, Vlatka Mičetić Stanković, Luka Čorak, Đurđica Ugarković, Albrecht Komarek. Contribution to molecular systematics of water scavenger beetles (Hydrophilidae, Coleoptera). „Journal of Zoological Systematics and Evolutionary Research”. 51, s. 165–171, 2013. DOI: 10.1111/jzs.12013.
- 1 2 Ladislav Bocak, Christopher Barton, Alex Crampton-Platt, Douglas Chesters, Dirk Ahrens, Alfried Vogler. Building the Coleoptera tree-of-life for >8000 species: Composition of public DNA data and fit with Linnaean classification. „Systematic Entomology”. 39 (1), s. 97-110, 2014. DOI: 10.1111/syen.12037.
- 1 2 D.D. McKenna, A.L. Wild, K. Kanda, C.L. Bellamy, R.G. Beutel, M.S. Caterino, C.W. Farnum, D.C. Hawks, M.A. Ivie, M.L. Jameson, R.A.B. Leschen, A.E. Marvaldi, J.V. McHugh, A.F. Newton, J.A. Robertson, M.K. Thayer, M.F. Whiting, J.F. Lawrence, A. Slipinski, D.R. Maddison, B.D. Farrell. The beetle tree of life reveals that Coleoptera survived end-Permian mass extinction to diversify during the Cretaceous terrestrial revolution. „Systematic Entomology”. 40 (4), s. 835-880, 2015. The Royal Entomological Society. DOI: 10.1111/syen.12132.
- 1 2 R.G. Beutel. Phylogenetic analysis of Hydrophiloidea based on characters of the head of adults and larvae. „Koleopterologische Rundschau”. 64, s. 103–131, 1994.
- ↑ Toby Hunt, Johannes Bergsten, Zuzana Levkanicova, Anna Papadopoulou, Oliver St John, Ruth Wild, Peter M Hammond, Dirk Ahrens, Michael Balke, Michael S Caterino, Jesús Gómez-Zurita, Ignacio Ribera, Timothy G Barraclough, Milada Bocakova, Ladislav Bocak, Alfried P Vogler. A comprehensive phylogeny of beetles reveals the evolutionary origins of a superradiation. „Science”. 318 (5858), s. 1913-1916, 2007. DOI: 10.1126/science.1146954.
- 1 2 3 Fikáček Martin, Prokin Alexander, Angus Robert B., Ponomarenko Alexander, Yue Yanli, Ren Dong, Prokop Jakub. Revision of Mesozoic fossils of the helophorid lineage of the superfamily Hydrophiloidea (Coleoptera: Polyphaga). „Acta Entomologica Musei Nationalis Pragae”. 52 (1), s. 89-127, 2012. ISSN 0374-1036.
- ↑ Chenyang Cai, Erik Tihelka, Mattia Giacomelli, John F. Lawrence, Adam Ślipiński, Robin Kundrata, Shûhei Yamamoto, Margaret K. Thayer, Alfred F. Newton , Richard A. B. Leschen, Matthew L. Gimmel, Liang Lü, Michael S. Engel, Patrice Bouchard, Diying Huang, Davide Pisani, Philip C. J. Donoghue. Integrated phylogenomics and fossil data illuminate the evolution of beetles. „Royal Society Open Science”. 9 (3), 2022. DOI: 10.1098/rsos.211771. ISSN 2054-5703.
- 1 2 3 Martin Fikáček, Alexander Prokin, Evgeny Yan, Yanli Yue, Bo Wang, Dong Ren, Robert Beattie. Original Article Modern hydrophilid clades present and widespread in the Late Jurassic and Early Cretaceous (Coleoptera: Hydrophiloidea: Hydrophilidae). „Zoological Journal of the Linnean Society”. 170 (4), s. 710-734, 2014. The Linnean Society of London. DOI: 10.1111/zoj.12114.