Oogamia - typ rozmnażania polegający na łączeniu się ruchliwej gamety męskiej - plemnika z nieruchliwą gametą żeńską - komórką jajową.
Anizogamia - typ rozmnażania polegający na łączeniu się dwóch gamet mających wici, ale różniących się wielkością wynikającą z różnej ilości materiału zapasowego - anizogamia żeńska posiada więcej materiału niż męska.
Izogamia - typ rozmnażania polegający na łączeniu się dwóch gamet jednakowych pod względem wielkości i ilości materiału zapasowego, zaopatrzonych w wici.
BRUNATNICE ZIELENICE KRASNOROSTY
1.WĘGLOWODANY (laminaryna i mannitol) tłuszcze. 1.SKROBIA 1.SKROBIA KRASNOROSTOWA, TŁUSZCZE
2. FUKOKSANTYNA, CHLOROFIL a, cKAROTEN.Główny ksantofil to fukoksantyna. 2. CHLOROFIL, KAROTENKSANTOFIL. CHLOROFIL a, b Główny ksantofil to luteina i zeaksantyna. 2. FIKOERYTRYNA FIKOCYJANINA CHLOROFIL a, dKAROTEN. Główny ksantofil to luteina i zeaksantyna oraz fikobiliny.
1. PRODUKTY FOTOSYNTEZY.2.BARWNIKI FOTOSYNTETYCZNE.
BRUNATNICE
Rozmnażają się bezpłciowo (przez zoospory i aplanospory) oraz płciowo sposobami oogami, anizogamii, izogamii.
KRASNOROSTY
Rozmnażają się poprzez przemianę pokoleń gametofitu (występują narządy płciowe, lęgnie zwane też karpogonami i plemnie tzw.spermatangium) , sporofitu.
ZIELENICE
Rozmnażają się bezpłciowo przez podział komórek, fragmentację plech lub wytwarzanie spor: ruchliwych zoospor i nieruchliwych aplanospor.
Płciowo rozmnażają się sposobem izogamii, anizogamii, oogami, niektóre również poprzez przemianę pokoleń.
ROZMNAŻANIE BRUNATNIC (na przykładzie morszczynu).
Morszczyn rozmnaża się wegetatywnie - przez fragmentację plechy oraz płciowo. W okresie rozmnażania płciowego na wierzchołkach rozgałęzień plechy rozwijają się komórki jajowe w lęgniach i plemniki w plemniach. Zależnie od gatunku morszczynu komórki rozrodcze obu płci powstają na jednej lub na dwóch różnych, rozdzielnopłciowych roślinach, zaopatrzone w wici plemniki oraz bezwiciowe komórki jajowe wydostają się z plemni i lęgni: zapłodnienie następuje w wodzie poza rośliną macierzystą. Zygota opada na dno dając początek nowej roślinie.
ROZMNAŻANIE KRASNOROSTÓW
W przemianie pokoleń charakterystyczny jest rozdział fazy diploidalnej na dwa pokolenia. Po wytworzeniu zygoty na gametoficie wyrasta pierwsze pokolenie diploidalne - karposporofit. Wytwarza on specyficznie, diploidalne zarodniki - karpospory. Z tych zarodników wyrasta drugie pokolenie diploidalne - tetrasporofit na którym powstają już klasyczne haploidalne zarodniki zwane tetrasporofitami.Z tetraspor wyrasta pokolenie haploidalne - gametofit.
ROZMNAŻANIE ZIELENIC
Rozmnażanie na drodze przemiany pokoleń. Wyróżnia się pokolenie bezpłciowe - diploidalny sporofit i pokolenie płciowe - haploidalny gametofit.Z haploidalnego zarodnika wyrasta gametofit, na którym tworzą się organa rozmnażania płciowego: lęgnie (lub rodnie) i plemnie. W nich powstają komórki płciowe odpowiednio żeńskie i męskie. Po zapłodnieniu czyli połączeniu się haploidalnych komórek płciowych (żeńskiej i męskiej) , powstaje diploidalna zygota - pierwsza komórka sporofitu. Na wyrosłym z zygoty sporoficie tworzą się zarodnie, w których zachodzi mejoza, prowadząca do wytworzenia haploidalnych zarodników, pierwszych komórek gametofitu. Jeśli gametofit i sporofit nie różnią się wyglądem miedzy sobą to mówimy o izomorficznej przemianie pokoleń, częściej jednak występuje heteromorficzna przemiana pokoleń czyli z zaznaczeniem dominacji gametofitu lub sporofitu.
Cykl haploidalny - występuje u glonów, których postać wegetatywna jest haploidalna.Stadium diploidalne reprezentuje tylko zygota i jej forma przetrwalnikowa. Mejoza zaś jest pierwszym podziałem podczas jej kiełkowania, a powstające komórki nie mają identycznego składu genetycznego i dają początek różniącym się genetycznie osobnikom.
Jeżeli komórkom powstającym w wyniku mejozy są zarodniki (spory) to nazywane są one mejosporami, a mejoza jest określana jako sporogeniczna.
Plemnie - anterydia
Lęgnie - oogonia
Wielokomórkowe lęgnie to rodnie lub archegonia.
MCHY (MUSCI).
Rośliny jednopienne - gdy rodnie i plemnie powstają na jednej roślinie.
Rośliny dwupienne - gdy rodnie i plemnie powstają na różnych osobnikach.
ROZMNAŻANIE MCHÓW (ZNACZNIE SKRÓCONE).
W cyklu życiowym mchów z zarodnika wyrasta stadium młodociane gametofitu - splątek.Z jednego splątka wyrasta następnie zwykle kilka dojrzałych gametofitów.Gametofit ma postać ulistnionej łodyżki. Listki zbudowane są z jednej warstwy komórek jedynie nerw przebiegający przez środek listka, jest wielowarstwowy. Mchy posiadają wielokomórkowe chwytniki. Zapłodnienie musi odbywać się w wodzie, najczęściej w kropli wody, deszczu. Sporofit zbudowany jest ze stopy, zwykle dużej sety i zarodni, zawierającej w środku charakterystyczną kolumienkę. Zarodnia okryta jest z zewnątrz czapeczką, która jest pozostałością po rozerwanej rodni.
Gametofit mszaków rozwija się z haploidalnego zarodnika.U mchów zarodnik kiełkuje w tzw. splątek - nitkowatą nietrwałą plechę, będącą stadium młodocianym gametofitu. Następnie z formujących się na splątku pączków rozwijają się ulistnione, zielone łodyżki będące właściwym gametofitem. Komórki gametofitu wykazują pewne zróznicowanie tkankowe, przy czym wszystkie tkanki stałe tworzące to pokolenie należy traktować jako odmiany lub modyfikacje rkanki miękiszowej. Na szczególną uwagę zasługują komórki przewodzące wodę i sole mineralne - tzw. Hydroidy oraz komórki przewodzące asymilaty - leptoidy.
PAPROTNIKI (PTERIDOPHYTA).
PAPROCIE (FILICINAE).
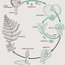
Rozmnażanie paproci. Następstwo pokoleń.
U paproci okazałe rośliny o złożonej budowie i funkcjach to właśnie sporofity - pokolenie diploidalne. Natomiast gametofit jest niepozorny i żyje krótko.
Sporofit paproci wytwarza zarodnie. Są one umieszczone na liściach, przeważnie na ich spodniej części. U niektórych paproci zarodnie powstają na specjalnie do tego celu przeznaczonych liściach zarodnionośnych tzw. Sporofilach.
Sporofile są wyłączone z funkcji asymilacyjnej. Na liściach naręcznicy samczej zarodnie występują w skupieniach tzw. Kupkach, widocznych jako brązowe plamki na spodzie liścia. Początkowo kupki przykryte są błoną zwana zawijką. W czasie dojrzewania zarodników zawijka wysycha i odsłania zarodnie. Mają one postać woreczków osadzonych na trzonkach. Każda zarodnia ma swoiste urządzenie otwierające - opasający ją pierścień komórek o odpowiednio zgrubiałych ścianach. Podczas suszy pierścień ten prostuje się, rozrywając ścianę zarodni i uwalniając zarodniki. Zarodniki podobnie jak u mchów są haploidalne; powstają w wyniku mejotycznych podziałów komórek macierzystych. Zarodnik zatem jest pierwszym stadium gametofitu. Kiełkujący zarodnik daje początek również haploidalnemu przedroślu. Przedrośle jest małym zielonym tworem sercowatego kształtu, o średnicy nie większej niż 1 cm. Związek z podłożem utrzymuje za pomocą nitkowatych chwytników. Na dolnej powierzchni przedrośla powstają rodnie z komórkami jajowymi i plemnie z plemnikami.
Plemniki są orzęsione zapłodnienie odbywa się w wodzie pokrywającej przedrośle. Rozmnażanie płciowe paproci, jest jednym momentem nawiązującym do wodnego trybu życia przodków tych roślin. W wyniku zapłodnienia komórki jajowej powstaje diploidalna zygota - zaczątek stadium sporofitowego. Rozrastający się młody sporofit początkowo czerpie substancje odżywcze ze stopniowo obumierającego przedrośla (gametofitu). Z chwilą ukorzenienia się rośliny i rozwinięcia się pierwszych liści sporofit usamodzielnia się , a przedrośle całkowicie zanika. Cykl następstwa pokoleń zaczyna się od nowa.
Następstwo pokoleń paproci jest wyrazem pełniejszego niż u mchów przystosowania do życia lądowego. U paproci obserwuje się przewagę sporofitu - pokolenia związanego z lądem, nad gametofitem - pokoleniem związanym z wodą. Dzięki temu paprocie mogły osiągnąć w warunkach lądowych znacznie wyższy poziom organizacji aniżeli mchy. Sporofit mchów nie styka się bezpośrednio z podłożem i nie ma możliwości rozrośnięcia się w samodzielną roślinę; gametofit również jest niepozorny - nie może wyrosnąć zbyt wysoko nad wilgotne podłoże ze względu na sposób rozmnażania związany z wodą.
Przeczytaj podobne teksty
Czas czytania: 7 minut
Ciekawostki ze świata
Treść zweryfikowana i sprawdzona
Zgodnie z misją, Redakcja serwisu dokłada wszelkich starań, aby dostarczać rzetelne i sprawdzone treści edukacyjne.
Dodatkowe oznaczenie "Sprawdzona treść" wskazuje, że dany materiał został zweryfikowany przez Redakcję lub ekspertów współpracujących z serwisem.
Weryfikacja tekstu odbywa się na następujących poziomach:
1. Sprawdzenie tekstu pod kątem ewentualnych błędów ortograficznych, stylistycznych, interpunkcyjnych lub innych.
2. Weryfikacja tekstu pod kątem błędów rzeczowych.
3. Sprawdzenie materiału pod kątem ich aktualności.
Dodatkowe oznaczenie "Sprawdzona treść" wskazuje, że dany materiał został zweryfikowany przez Redakcję lub ekspertów współpracujących z serwisem.
Weryfikacja tekstu odbywa się na następujących poziomach:
1. Sprawdzenie tekstu pod kątem ewentualnych błędów ortograficznych, stylistycznych, interpunkcyjnych lub innych.
2. Weryfikacja tekstu pod kątem błędów rzeczowych.
3. Sprawdzenie materiału pod kątem ich aktualności.