Oddychanie tlenowe zachodzi w trzech etapach:
glikoliza,
cykl kwasu cytrynowego (cykl Krebsa),
łańcuch oddechowy.
Glikoza jest ciągiem reakcji przekształcających glukozę w pirogronian bez udziału tlenu. Zachodzi w cytoplazmie. U organizmów tlenowych jest wstępem do cyklu Krebsa i łańcucha transportującego elektrony. Natomiast u organizmów beztlenowych oraz u tlenowych w warunkach deficytu tlenu (np. w mięśniach) jest zasadniczym sposobem pozyskiwania energii. Podczas glikolizy 1 cząsteczka glukozy zostaje rozłożona na 2 cząsteczki trójwęglowego związku – pirogronianu.
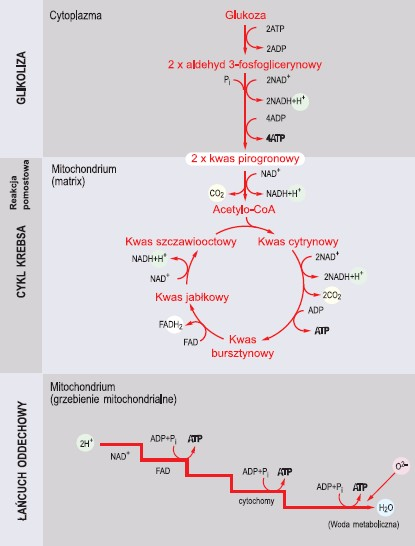
Główne etapy oddychania tlenowego (wg Wiśniewski, 1994, zmienione)
W początkowych fazach procesu następuje dwukrotna fosforylacja, co wymaga zużycia 2 cząsteczek ATP. Później energia chemiczna uwalniana podczas przekształceń trójwęglowych produktów pośrednich (aldehydu 3-fosfoglicerynowego) pozwala na przyłączenie kolejnych reszt fosforanowych, po jednej do każdej cząsteczki produktu. Przeniesienie wszystkich reszt fosforanowych z ich wiązaniami wysoko energetycznymi na ADP umożliwia syntezę, na drodze fosforylacji substratowej, 4 cząsteczek ATP. Zysk glikolizy stanowią 2 cząsteczki ATP oraz 2 cząsteczki zredukowanego NADH, który powstaje dzięki dehydrogenacji aldehydu 3-fosfoglicerynowego. Aby proces mógł zachodzić w sposób ciągły, NADH+H+ musi ulec z powrotem utlenieniu do NAD+, co w warunkach tlenowych zachodzi w łańcuchu oddechowym, albo w inny sposób podczas oddychania beztlenowego.
W oddychaniu tlenowym wytworzony w glikolizie kwas pirogronowy w tzw. reakcji pomostowej zostaje przekształcony w acetylo-CoA (dekarboksylacja z równoczesnym utlenieniem dwuwęglowego produktu i przyłączeniem do niego koenzymu A). W reakcji tej odłączają się dwa atomy wodoru (2 protony i 2 elektrony), które przejmuje NAD+, ulegając redukcji do NADH+H+. Proces zachodzi w matrix mitochondrialnej.
Następny etap to cykl kwasu cytrynowego, zachodzący także w matrix mitochondriów, polega na utlenieniu acetylo-CoA do dwutlenku węgla oraz atomów wodoru (w postaci elektronów o wysokiej energii i protonów). W cyklu Krebsa grupy acetylowe acetylo-CoA nie są utleniane bezpośrednio, ale przenoszone z acetylo-CoA do czterowęglowej cząsteczki kwasu szczawiooctowego. Powstaje wówczas sześciowęglowy kwas cytrynowy, który ulega kolejnym przemianom zmierzającym do odtworzenia kwasu szczawiooctowego. Wśród reakcji cyklu najistotniejsze są dwie dekarboksylacje, prowadzące do powstania 2 cząsteczek CO2, oraz cztery dehydrogenacje, które uwalniają z produktów pośrednich cyklu protony i elektrony przyłączane następnie do 3 cząsteczek NAD+ (redukcja NAD+ do NADH+H+) oraz l cząsteczki FAD (redukcja FAD do FADH2). NADH+H+ oraz FADH2 zostają później przekazane do łańcucha oddechowego. Dodatkowym zyskiem cyklu Krebsa są 2 cząsteczki GTP (guanozynotrifosforanu) równoważne 2 cząsteczkom ATP, powstające podczas fosforylacji substratowej. Cykl Krebsa sam nie zużywa tlenu, ale potrzebuje go do odtworzenia NAD+ i FAD, co następuje podczas przekazywania elektronów na cząsteczkę O2 w łańcuchu oddechowym przez NADH+H+ oraz FADH2.
Znaczenie cyklu Krebsa dla komórki polega na tym, że włączany do niego acetylo-CoA pochodzi zarówno z przemian węglowodanów, jak i tłuszczów oraz białek. Ponadto wiele metabolitów pośrednich cyklu jest wykorzystywana przez organizm do różnych syntez.
Ostatnim etapem oddychania tlenowego jest łańcuch oddechowy, który składa się z przenośników elektronów, zlokalizowanych w wewnętrznej błonie mitochondriów. Przenośnikami tymi są głównie białka transbłonowe, zawierające ściśle z nimi związane metale o właściwościach oksydoredukcyjnych. Część białek zgrupowana jest w trzech enzymatycznych kompleksach oddechowych (dehydrogenaza NADH, cytochromy, oksydaza cytochromowa). W łańcuchu oddechowym przenośniki elektronów ułożone są według wzrastającego potencjału oksydoredukcyjnego, co umożliwia przechwytywanie elektronów przez kolejne przenośniki obdarzone coraz większym do nich powinowactwem. Znajdujący się przy końcu łańcucha O2 wyłapuje elektrony i równocześnie przyłącza obecne w matrix H+. W ten sposób powstaje cząsteczka H2O. W każdym takim przeniesieniu elektronu uwalniana jest energia służąca do transportu protonów z matrix do przestrzeni między błoną zewnętrzną a wewnętrzną mitochondrium. Efektem przepompowania protonów przez błonę jest różnica stężeń H+ po obu stronach błony. Wyrównanie gradientu protonowego następuje dzięki powrotowi H+ do matrix przez kompleks enzymatyczny, zwany syntazą ATP, który wykorzystuje energię przepływających protonów do syntezy ATP. Zatem do wytworzenia ATP zużywana jest energia wyzwolona podczas przepływu elektronów przez łańcuch oddechowy, gdyż to ona warunkuje ruch protonów. Ten sposób fosforylowania ADP do ATP zwany jest fosforylacją oksydacyjną. Łączny zysk z transportu elektronów w łańcuchu oddechowym wynosi około 32 cząsteczki ATP. Związek między transportem elektronów, pompowaniem protonów i syntezą ATP określany jest mianem mechanizmu sprzężenia chemiosmotycznego.