| Rhyniognatha | |
| Tillyard, 1928 | |
| Okres istnienia: dewon wczesny | |
 | |
| Systematyka | |
| Domena | |
|---|---|
| Królestwo | |
| Typ | |
| Rodzaj |
Rhyniognatha |
| Typ nomenklatoryczny | |
|
Rhyniognatha hirsti Tillyard, 1928 | |
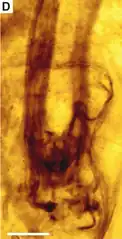
Rhyniognatha – wymarły rodzaj stawonogów, obejmujący tylko jeden opisany gatunek: Rhyniognatha hirsti. Znany jest wyłącznie z pojedynczej, bardzo niekompletnej skamieniałości głowy, obejmującej głównie elementy narządów gębowych. Znaleziona została w szkockim Rhynie Chert i pochodzi z pragu (dewon wczesny, około 396–407 mln lat temu)[1]. Przynależność rodzaju do gromady jest niejasna[2]. Opisywany był trzykrotnie: w 1926 uznany został za owada bez przypisywania do rzędu[3], w 2004 za najstarszego przedstawiciela owadów uskrzydlonych, nawiązującego budową do Metapterygota[1], a w 2017 za z większym prawdopodobieństwem wija z gromady pareczników[2].
Historia odkrycia i badań
Próbka skały zawierająca okaz Rhyniognatha została pozyskana w Rhynie Chert w 1919 przez R.W. Crana i dostarczona S. Hirstowi, S. Maulikowi i D.J. Scourfieldowi. W 1926 S. Hirst i S. Maulik na podstawie znajdujących się w niej skamieniałości opisali nowy gatunek i rodzaj stawonoga: Rhyniella praecursor, który współcześnie zaliczany jest do skoczogonków. Do owego rodzaju zaliczyli cztery mniej lub lepiej zachowane skamieniałości głów oraz jedną skamieniałość „szczęk”[4]. Skamieniałości te zostały ponownie przebadane przez R.J. Tillyarda. W publikacji z 1928 wyznaczył najlepiej zachowaną głowę holotypem Rhyniella praecursor, a pozostałe trzy głowy paratypami. Natomiast parę „szczęk” Tillyard rozpoznał jako żuwaczki owada i opisał jako nowy gatunek i rodzaj: Rhyniognatha hirsti, nadając epitet gatunkowy na cześć S. Hirsta[3].
Przez kolejne lata interpretacje okazu podawano za Tillyardem[5][1], uznawano go za okaz o niejasnym pochodzeniu[6] lub też zupełnie go ignorowano[7]. W 2002 A.P. Rasnicyn i D.L. Quicke zasugerowali, że stawonóg ten był raczej wijem niż owadem[8], ale pierwszego ponownego przebadania okazu dokonali M.S. Engel i D.A. Grimaldi za pomocą mikroskopu złożonego. Ich publikacja ukazała się w 2004 na łamach Nature. Zinterpretowali oni żuwaczki jako mające dwa funkcjonalne połączenia stawowe (dykondyliczne) i wyraźny podział krawędzi wewnętrznej na tnącą i molarną. Na tej podstawie zaliczyli oni Rhyniognatha do owadów uskrzydlonych, wskazując na silne podobieństwo jej żuwaczek do tych u Metapterygota (linii ewolucyjnej obejmującej wszystkie owady uskrzydlone oprócz jętek)[1]. Miało to duże implikacje dla wiedzy o ewolucji owadów, gdyż wówczas za najstarsze znane szczątki właściwych owadów (jawnoszczękich) uważano pochodzące sprzed około 390 mln lat fragmenty pancerzyków przerzutek[9][1] lub pochodzące sprzed około 379 mln lat fragmenty oskórka rybików lub przerzutek[10][1] – w obu przypadkach były to starsze ewolucyjnie owady bezskrzydłe (szczeciogonki). Zapis kopalny jako początek linii owadów uskrzydlonych wskazywał wówczas na dewon późny lub wczesny karbon. Rozpoznanie w Rhyniognatha owada uskrzydlonego oznaczało cofnięcie ich początku co najmniej do dewonu wczesnego. Engel i Grimaldi wskazali na zgodność takiego poglądu z wynikami Gaunta i Miliesa z 2002, którzy za pomocą zegaru molekularnego datowali pojawienie się owadów na 434 mln lat temu (wczesny sylur), a nowoskrzydłych na 387 mln lat temu (dewon środkowy)[1][11][12]. Interpretacja Engela i Grimaldiego sprawiła, że Rhyniognatha była wykorzystywana do kalibracji zegarów molekularnych w późniejszych badaniach[2], np. Warnocka i innych z 2011[13], Misofa i innych z 2014[14] oraz Wolfego i innych z 2016[15].
Ponownego przebadania skamieniałości Rhyniognatha dokonali C. Haug i J. Haug, wykorzystując stacking i obrazowanie 3D. Nie zidentyfikowali oni dwóch połączeń stawowych żuwaczki, natomiast rozpoznali w skamieniałości fragment puszki głowowej (egzoszkieletu głowy) niedostrzeżony przez Engela i Grimaldiego. Parzyste struktury opisane przez Engela i Grimaldiego jako apodemy zinterpretowali z większym prawdopodobieństwem jako gruczoły żuwaczkowe. Według wyników ich pracy pewne przypisanie Rhyniognatha do konkretnej gromady stawonogów nie jest możliwe, jednak z większym prawdopodobieństwem jest to parecznik niż owad[2].
Opis
Jedyna znana skamieniałość obejmuje kilka w różnym stopniu połączonych fragmentów oskórka, z których te należące niewątpliwie do Rhyniognatha, zidentyfikowane zostały jako części sześciu narządów głowy[2]. Najbardziej rzucającym się w oczy elementem są parzyste żuwaczki[11][1][2], z których prawa ma długość 0,17 mm[3]. Były one silnie zesklerotyzowane, krótkie i trójkątne w zarysie. W interpretacji Engela i Grimaldiego ich krawędzie wewnętrzne były wyraźnie zróżnicowane na wierzchołkową część tnącą (incisivus) i nasadową część molarną (mola), przy czym krawędź tnąca zaopatrzona była w trzy duże i dwa małe ząbki, a część molarna pozbawiona pędzla szczecinek[1]. Według Haug i Hauga zróżnicowanie na część tnącą i molarną jest słabiej zaznaczone, a ta pierwsza ma tylko trzy duże zęby, zaś pozostałe dwa nie są wyraźnie widoczne i równie dobrze mogą być częścią innego narządu gębowego[2]. Największe różnice w interpretacji dotyczą nasadowej (dosiebnej) powierzchni żuwaczek. Według Grimaldiego i Engela jej kąt wewnętrzny zaopatrzony jest w panewkę stawową (acetabulum) przedniego kłykcia (condylus anterioris), a kąt zewnętrzny tworzy tylny kłykieć (condylus posterioris). Taka budowa pozwalałaby zaklasyfikować Rhyniognatha nie tylko jako owada, ale także jako dość zaawansowanego ewolucyjnie owada uskrzydlonego z kladu Metapterygota. Dwukłykciowe połączenie żuwaczki pozwalałoby odróżnić Rhyniognatha od skrytoszczękich i przerzutek, jej kształt i wyodrębnienie pozbawionej pędzelka krawędzi molarnej od rybików, a kształt i obecność dobrze wykształconego, funkcjonalnego przedniego połączenia stawowego od jętek[1]. Badania Haug i Hauga nie potwierdziły jednak obecności panewek stawowych w nasadowych częściach żuwaczek. Według tych autorów ich rejon dosiebny jest trudny w interpretacji i nie da się orzec, czy żuwaczki miały połączenie dwukłykciowe czy nie[2].
Kolejnym elementem parzystym aparatu gębowego w skamieniałości są wyrostki wystające przed żuwaczki i biorące początek nieco poniżej nich[2]. W interpretacji Engela i Grimaldiego ich wierzchołki są rozwidlone i autorzy najpierw opisali je jako żuwki wewnętrzne[1], a w późniejszej publikacji mniej precyzyjnie jako elementy szczęk[11]. Według Haug i Hauga wierzchołki tych struktur nie są rozdwojone, a ich powierzchnia jest słabiej zesklerotyzowana i miejscami pomarszczona. Obrazowanie 3D wskazuje, że narządy te są bardzo wydłużone i ostro zakrzywione do wewnątrz, stąd zinterpretowane zostały przez tych autorów jako głaszczki[2]. W otoczeniu żuwaczek obecna jest również czapkowata struktura, niepodpisana przez Engela i Grimaldiego, natomiast według Haug i Hauga najprawdopodobniej stanowiąca wargę górną lub część kompleksu wargowo-nadustkowego. Ponadto Haug i Haug zaobserwowali parę grzebieniastych narządów wystających spod żuwaczek i w większej części przez nie nakrytych, które uznali za najprawdopodobniej szczęki pierwszej pary[2].
Od żuwaczek ku tyłowi ciągnie się para podłużnych struktur o kształcie łyżki do butów. Ich szerokość odpowiada mniej więcej szerokości żuwaczek, natomiast długość jest ośmiokrotnie większa[2]. Według publikacji Engela i Grimaldiego z 2004 stanowią one apodemy[1]. W publikacji z 2005 autorzy Ci opisali je natomiast jako fragmenty puszki głowowej[11]. Taka interpretacja została odrzucona przez Haug i Hauga. Autorzy ci zaobserwowali fragment interpretowany jako należący do właściwej puszki głowowej, niedostrzeżony przez Engela i Grimaldiego. W przedniej części otacza on w pewnej odległości żuwaczki, biegnąc powyżej nich, następnie biegnie lewym bokiem, wypuszczając w części środkowo-grzbietowej krótkie beleczki w kierunku dośrodkowym, po czym skręca w prawo i przebiega nad parą omawianych podłużnych struktur. Prawa część puszki głowowej jest uznawana za zniszczoną i stąd niekompletną, natomiast wspomniane beleczki jako pasma szkieletu wokół otworów na oczy i czułki. Taka budowa puszki głowowej wskazuje na przedstawiciela owadów lub wijów, wykluczając równonogi. Jednak fakt, iż omawiane podłużne struktury sięgają daleko poza obrys puszki, wskazuje według Haug i Hauga bardziej na wije, gdyż u owadów puszka głowowa jest z tyłu silniej zamknięta, a przez mały otwór potyliczny utrudnia przechodzenie do tułowia tak dużych elementów. Haug i Haug zaznaczają, że mogą one stanowić mięśnie lub apodemy, ale szczególną uwagę przywiązują do interpretacji ich jako ektodermalne gruczoły żuwaczkowe pareczników. Na korzyść tej hipotezy świadczy umiejscowienie owych narządów mniej więcej na poziomie żuwaczek (ich przednie części mogą wręcz wchodzić do wnętrza żuwaczek). Ponadto ich powierzchnie wyglądają na pomarszczone i wskutek tego słabiej zesklerotyzowane, co mogłoby odpowiadać ektodermalnym wgłobieniom obserwowanym w takich gruczołach. Według Haug i Hauga możliwe jest, że R. hirsti jest w rzeczywistości młodocianą formą Crussolum – rodzaju wczesnego parecznika z rzędu przetarcznikokształtnych, opisanego również z Rhynie Chert na podstawie pazura jadowego i pojedynczych odnóży[2].
Przypisy
- 1 2 3 4 5 6 7 8 9 10 11 12 Michael S. Engel, David A. Grimaldi. New light shed on the oldest insect. „Nature”. 427 (6975), s. 627-630, 2004. DOI: 10.1038/nature02291.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 Carolin Haug, Joachim T. Haug. The presumed oldest flying insect: more likely a myriapod?. „PeerJ”. 5, 2017. DOI: 10.7717/peerj.3402.
- 1 2 3 Robert John Tillyard. Some remarks on the Devonian fossil insects from the Rhynie chert beds, Old Red Sandstone. „Transactions of the Royal Entomological Society of London”. 76 (1), s. 65-71, 1928.
- ↑ Arthur Stanley Hirst, Samarendra Nath Maulik. On some arthropod remains from the Rhynie chert (Old Red Sandstone). „Geological Magazine”. 63 (2), s. 69-71, 1926.
- ↑ W. Hennig: Insect Phylogeny. New York: John Wiley & Sons, 1981.
- ↑ D.J. Scourfield. The oldest known fossil insect. „Nature”. 145, s. 799–801, 1940.
- ↑ F.M. Carpenter, R.L. Kaesler: Treatise on Invertebrate Paleontology, Part R: Arthopoda 4: Superclass Hexapoda. Boulder, Colorado: Geol. Soc. Am, 1992.
- ↑ Aleksandr P. Rasnicyn, Donald L. Quicke: History of insects. Dordrecht: Kluwer Academic Publishers, 2002.
- ↑ C.C. Labandeira, B.S. Beall, F.M. Hueber. Early insect diversification: Evidence from a Lower Devonian bristletail from Quebec. „Science”. 242, s. 913–916, 1988.
- ↑ William A. Shear, P.M. Bonamo, J.D. Grierson, W.I. Rolfe, E.L. Smith, R.A. Norton. Early land animals in North America: Evidence from Devonian age arthropods from Gilboa, New York. „Science”. 224, s. 492–494, 1984.
- 1 2 3 4 David A. Grimaldi, Michael S. Engel: Evolution of the insects. Cambridge: Cambridge University Press, 2005.
- ↑ M.W. Gaunt, M.A. Miles. An insect molecular clock dates the origin of the insects and accords with palaeontological and biogeographic landmarks. „Mol. Biol. Evol.”. 19, s. 748–761, 2002.
- ↑ R.C. Warnock, Z. Yang, P.C. Donoghue. Exploring uncertainty in the calibration of the molecular clock. „Biology Letters”, 2011.
- ↑ Bernhard Misof i inni, Phylogenomics resolves the timing and pattern of insect evolution, „Science”, 346, 2014, s. 763-767, DOI: 10.1126/science.1257570, PMID: 25378627, OCLC 5694389053.
- ↑ J.M. Wolfe, A.C. Daley, D.A. Legg, G.D. Edgecombe. Fossil calibrations for the arthropod Tree of Life. „Earth-Science Reviews”. 160, s. 43-110, 2016.