| Lambeosaurus | |||
| Parks, 1923 | |||
| Okres istnienia: 76–75 mln lat temu | |||
 | |||
| Systematyka | |||
| Domena | |||
|---|---|---|---|
| Królestwo | |||
| Typ | |||
| Podtyp | |||
| Gromada | |||
| Podgromada | |||
| Nadrząd | |||
| Rząd | |||
| Podrząd | |||
| Infrarząd | |||
| Nadrodzina | |||
| Rodzina | |||
| Podrodzina | |||
| Plemię | |||
| Rodzaj |
lambeozaur | ||
| Gatunki | |||
| |||
Lambeozaur (Lambeosaurus, „jaszczur Lambe'a”) − rodzaj dinozaura kaczodziobego, żyjącego od 76 do 75 milionów lat temu, w kampanie, w kredzie późnej, w Ameryce Północnej. Ten dwu- lub czworonożny roślinożerca znany jest z charakterystycznych wydrążonych tworów na głowie, u najlepiej poznanego gatunku przypominających toporek. Wyróżniono kilka prawdopodobnych gatunków z Alberty w Kanadzie i Montany w Stanach Zjednoczonych, jednak tylko dwa gatunki kanadyjskie są obecnie dobrze poznane.
Lambeosaurus został opisany przez Williama Parksa późno: w 1923, ponad 20 lat po zbadaniu pierwszych szczątków przez Lawrence'a Lambe'a. Rodzaj cechuje się skomplikowaną historią taksonomiczną, po części dlatego, że mniejsze grzebieniaste kaczodziobe, dzisiaj uważane za osobniki młodociane, umieszczano niegdyś w osobnych rodzajach i gatunkach. Obecnie zróżnicowane czaszki przypisywane gatunkowi typowemu L. lambei interpretowane są jako efekt różnego wieku zwierząt lub dymorfizmu płciowego. Lambeozaur spokrewniony był blisko z lepiej poznanym korytozaurem, znajdywanym w nieco starszych skałach, a także z mniej znanym hipakrozaurem czy olorotytanem. Wszystkie one dysponowały ciekawymi grzebieniami, którym przypisuje się funkcje społeczne: udział w generowaniu dźwięków i rozpoznawaniu się zwierząt.
Budowa

Lambeosaurus, w tym najlepiej poznany L. lambei, wykazywał znaczne podobieństwo do szeroko znanego korytozaura we wszystkim prócz struktur zdobiących głowę. W porównaniu z korytozaurzym grzebień lambeozaura kierował się bardziej do przodu, a wydrążone zatoki w jego środku znajdowały się w przedniej jego części, przebiegając wertykalnie[1]. Kolejną różnicę między rodzajami stanowi brak rozwidlonych wyrostków kości nosowej współtworzącej boczną część grzebienia. Jest to jedyna cecha różnicująca młode osobniki obydwu rodzajów, gdyż grzebienie przybierały swe charakterystyczne formy z wiekiem[2].
Budową Lambeosaurus przypominał inne hadrozaury. Potrafił więc poruszać się na dwóch lub na czterech kończynach, za czym przemawiają ślady spokrewnionych z nim zwierząt. Jego długi ogon usztywniały skostniałe ścięgna, chroniąc go przed opadaniem. Dłonie kończyły się czterema palcami, brakowało kciuka. Trzy kolejne palce (odpowiadające ludzkim palcom: wskazującemu, środkowemu i serdecznemu) łączyły się ze sobą, nosiły kopytka, co wspiera tezę o wspieraniu się gada na kończynach przednich. Ostatni palec (odpowiadający ludzkiemu palcu małemu), wolny, mógł służyć do manipulacji przedmiotami. Każda ze stóp miała tylko trzy centralne palce[3].
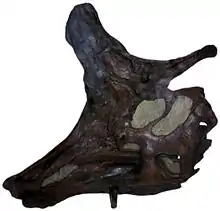
Najbardziej charakterystyczna cecha zwierzęcia, grzebień, różniła się u dwóch najlepiej poznanych gatunków. W przypadku L. lambei osiągał on u w pełni rozwiniętego osobnika kształt toporkowaty. U zwierząt uznawanych za samice był nieco krótszy i bardziej zaokrąglony[1]. Toporkowate ostrze wystawało do przodu naprzeciwko oczu, natomiast „uchwyt toporka” tworzył solidny kostny pręt wystający z tyłu czaszki. „Ostrze toporka” dzieliło się na dwie części: górna stanowiła cienki kostny grzebień wyrastający w trakcie ontogenezy stosunkowo późno, gdy zauropsyd był już prawie dorosły, dolna zawierała wydrążone przestrzenie stanowiące kontynuację jam nosowych[1]. U L. magnicristatus „uchwyt” był znacznie zredukowany, „ostrze” − dobrze rozwinięte[4], tworząc wysoki, wręcz nieprzystający grzebień przypominający fryzurę w stylu Madame Pompadour. U najlepiej zachowanego okazu grzebień ten uległ uszkodzeniu, pozostała jedynie jego przednia połowa[5]. Nieznany jest grzebień gorzej poznanego gatunku L. paucidens.
Gatunki kanadyjskie zdają się bardziej przypominać wymiarami korytozaura, co oznacza około 9,4 m długości[6]. Kilka okazów dostarczyło informacji o odciśniętych łuskach. Jeden z nich, dziś zaliczany do L. lambei, pokrywała cienka skóra o jednolitych, wielokątnych łuskach nie tworzących szczególnych rzędów na szyi, tułowiu i ogonie[7]. Podobny układ łusek znaleziono na szyi, przednich łapach i stopach okazu przypisywanego L. magnicristatus[5].
Systematyka
Lambeosaurus stanowi rodzaj typowy Lambeosaurinae, podrodziny hadrozaurów o wydrążonych grzebieniach na czaszce. Wśród lambeozaurynów jego bliskimi krewnymi są korytozaur i hipakrozaur, od których odróżnia się nieco budową grzebienia[3]. Pokrewieństwo pomiędzy tymi rodzajami dinozaurów jest trudne do ustalenia. Spotykane w internecie klasyfikacje czasami łączą te trzy rodzaje w plemię Lambeosaurini, nie zostało ono jednak dotychczas formalnie zdefiniowane. Zamiast niego niektóre klasyfikacje wymieniają równoważny klad Corythosaurini. Zalicza się tutaj najnowszy pogląd na filogenezę tej grupy zaprezentowany przez Davida Evansa i Roberta Reisza w ich redeskrypcji L. magnicristatus. Wedle tych badań Lambeosaurus stanowi takson siostrzany dla kladu tworzonego przez korytozaura, hipakrozaura i rosyjski rodzaj Olorotitan. Lambeosaurinae te razem z Nipponosaurus tworzą Corythosaurini[5].
Odkrycie i historia

Skomplikowana historia taksonomiczna Lambeosaurus zaczęła się w 1902, kiedy to Lawrence Lambe nazwał pozostałości kończyny i inne kości hadrozauryda z Alberty, oznakowane pierwotnie jako GSC 419, binominalną nazwą Trachodon marginatus[8]. W latach dziesiątych XX wieku paleontolodzy zaczęli znajdować lepszej jakości szczątki kaczodziobych w tych samych skałach, znanych obecnie jako późnokampańska (kreda górna) formacja Dinosaur Park. Lambe zaliczył do T. marginatus dwie nowe czaszki, bazując na nowych danych ukuł dla gatunku w 1914 nazwę rodzajową Stephanosaurus[9]. Niestety bardzo mało łączyło rzeczone czaszki z fragmentarycznym wcześniejszym materiałem określonym epitetem marginatus, wobec czego w 1923 William Parks zaproponował umieszczenie ich w nowym rodzaju i gatunku, w obu nazwach honorując Lambego i tworząc Lambeosaurus lambei (okaz typowy NMC 2869, pierwotnie GSC 2869)[10].
Nowe gatunki i Procheneosaurus
Nie zdając sobie z tego sprawy, wcześni poszukiwacze znajdywali już w Albercie szczątki młodocianych lambeozaurów. Te skamieliny niewielkich grzebieniastych kaczodziobych interpretowano jako odrębną linię hadrozaurów, podrodzinę Cheneosaurinae[11]. Pierwsze takie zwierzę nazwano Didanodon altidens na podstawie lewej szczęki GSC 1092 z formacji Dinosaur Park opisanej przez Henry'ego Osborna w 1902[12]. Została jednak umieszczona w taksonach Trachodon altidens przez Lambego[8], a nazwa Didanodon szybko wyszła z użycia. Nie wymieniają jej Richard Swann Lull i Nelda Wright we wpływowej monografii z 1942 traktującej o hadrozaurach Ameryki Północnej.

W 1920 William Diller Matthew użył nazwy rodzajowej Procheneosaurus bez epitetu gatunkowego w podpisie fotografii szkieletu z Amerykańskiego Muzeum Historii Naturalnej z formacji Dinosaur Park (AMNH 5340)[13]. Park uznawał taką procedurę i opis za nieadekwatne dla uznawanej za pewną nazwy i w związku z zaistniałą sytuacją ukuł nazwę Tetragonosaurus. W obrębie rodzaju umieścił gatunek typowy T. praeceps (bazując na ROM 3577) i drugi gatunek T. erectofrons (opierając się na ROM 3578) o małych czaszkach z formacji Dinosaur Park. Przypisał też szkielet Matthew znany jako Procheneosaurus gatunkowi T. praeceps[14]. Charles Mortram Sternberg poszedł w 1935 w tym samym kierunku, dodając nieco większego T. cranibrevis, bazując na GSC (obecnie NMC) 8633[15]. Użycie Tetragonosaurus zostało odrzucone przez Lulla na korzyść Procheneosaurus. W 1942 razem z Wrightem przenieśli gatunki Tetragonosaurus i, tymczasowo, Trachodon altidens, do Procheneosaurus z P. praeceps jako gatunkiem typowym[16]. Ich śladem podążano do 1975, kiedy Peter Dodson wykazał, że „Cheneosaurinae” stanowiły w rzeczywistości młodociane formy innych dinozaurów[1]. Procheneosaurus praeceps i altidens uznano za prawdopodobne młodsze synonimy dla Lambeosaurus lambei[3], choć oba wyprzedzały chronologicznie tę nazwę.
„Procheneosaurus” nie były jedynymi grzebieniastymi kaczodziobymi przestudiowanymi i nazwanymi na początku XX wieku. Stanowiło wtedy akceptowaną praktykę nadawanie nazw rodzajowych i gatunkowych formom uznawanym obecnie za przejaw zmienności osobniczej, spowodowanej wiekiem i płcią zwierzęcia, a także zniekształceniami podczas fosylizacji. W tym okresie wyróżniono więcej gatunków powiązanych z Lambeosaurus − wszystkie trzy opisano w 1935. Sternberg, w tej samej publikacji, co T. cranibrevis, nadał nazwy czaszkom i fragmentarycznym szkieletom GSC − obecnie NMC − 8705 L. magnicristatum (skorygowane na magnicristatus) i mniejszej czaszce GSC − obecnie NMC − 8703 L. clavinitialis, z mniej wydatnym grzebieniem i zredukowanym kolcem z tyłu[15]. Parks poparł gatunek Corythosaurus frontalis, oparty na czaszce GSC 5853 (obecnie ROM 869), różniący się grzebieniem innym od dobrze znanego, wysokiego, zaokrąglonego grzebienia innych korytozaurów − niskim i skierowanym w przód[17].
Zmiana poglądów
Po dużej aktywności na początku XX wieku przez wiele lat nie opisano ani jednego okazu. W 1964 John Ostrom zauważył, że starszy gatunek, nazwany przez Marsha Hadrosaurus paucidens, bazujący na USNM 5457, niekompletnej szczęce i kości łuskowej z formacji Judith River z hrabstwa Fergus w Montanie, stanowił prawdopodobnie okaz lambeozaura[18].
W 1975 Peter Dodson, badając zagadkę różnorodności gatunków Lambeosaurinae żyjących w tak krótkim okresie na małym obszarze, opublikował wyniki badania morfometrycznego, w którym zmierzył tuziny czaszek. Zauważył, że wiele gatunków bazuje na szczątkach, które lepiej interpretować jako pozostałości osobników młodocianych lub odmiennej płci. W przypadku Lambeosaurus odkrył, że L. clavinitialis był prawdopodobnie samicą L. lambei, natomiast Corythosaurus frontalis czy Procheneosaurus praeceps stanowiły prawdopodobnie osobniki młodociane tego gatunku. L. magnicristatus wyróżniał się dostatecznie, by dać podstawę dla umieszczenia w oddzielnym gatunku. Naukowiec uznał też Procheneosaurus cranibrevis i P. erectofrons za młode korytozaury[1]. Jednakże powtórne przestudiowanie szczątków Procheneosaurus/Tetragonosaurus pokazało, że klasyfikowanie ich w obrębie tego gatunku było mylne, a okaz typowy P. cranibrevis to młodociany lambeozaur, podczas gdy inne zaliczały się do Corythosaurus. Wnioski te opierają się o odmienne formy spojenia pomiędzy kością nosową i przysieczną[2].
Gatunki
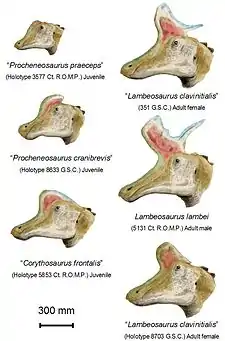
Obecnie za pewne uznaje się tylko dwa gatunki lambeozaura, trzeci jest szeroko akceptowany, czwarty zaś tylko czasami. L. lambei Parks, 1923 znany jest z co najmniej 17 osobników. Znaleziono 7 czaszek i niekompletnych szkieletów oraz około 10 izolowanych czaszek. L. clavinitialis Sternberg, 1935, Corythosaurus frontalis Parks, 1935 i Procheneosaurus praeceps Parks, 1931 uznaje się w najnowszych pracach za synonimy L. lambei[3]. Możliwe, że czaszka L. clavinitialis bez odchylającego się w tył wyrostka reprezentuje L. magnicristatus[19], choć opcję tą odrzucono w redeskrypcji L. magnicristatus z 2007[5]. Wiedza o L. magnicristatus Sternberg, 1935 pochodzi definitywnie tylko z dwóch okazów, w obu przypadkach czaszek. Niestety utracono większość połączonego stawowo szkieletu holotypowego. Wiele kości doznało rozległych uszkodzeń spowodowanych wodą w miejscu przechowywania i trzeba je było odrzucić jeszcze przed opisem. Stracono też inne części szkieletu. Szczątki te pochodzą z nieco młodszych skał, niż L. lambei[5]. Epitet gatunkowy pochodzi od łacińskich słów magnus (wielki) i cristatus (grzebieniasty). Odnosi się on do kostnego grzebienia zwierzęcia[20]. Ponadto Jack Horner zidentyfikował fragmentarycznie szczęki Lambeosaurinae z formacji Bearpaw z Montany jako prawdopodobnie należące do L. magnicristatus; jako pierwsze reprezentują one szczątki lambepoaurynów ze skał morskich[21]. Ogromny meksykański gatunek „L.” laticaudus Morris, 1981[22], znany z niepołączonych pozostałości kilku osobników, bywał przez część autorów uznawany za osobny gatunek lambeozaura[3], a przez innych za gatunek hipakrozaura[5][23]. Jego pewność zakwestionowali autorzy opisu Velafrons, uważający jego pozostałości za niediagnostyczne dla tego rodzaju[23]. Ostatecznie gatunek ten w 2012 r. ustanowiony został gatunkiem typowym odrębnego rodzaju Magnapaulia[24].
L. paucidens (Marsh, 1889) uznaje się za nomen dubium, a jego szczątki zalicza się wedle najnowszych poglądów do Hadrosaurus paucidens[3], choć co najmniej jeden autor, Donald F. Glut, uważa go za gatunek lambeozaura[25]. W tym przypadku epitet gatunkowy pochodzi od łacińskich pauci- (niewiele) i dens (ząb)[20]. Nieprawidłowości w związku z Procheneosaurus cranibrevis i jego identyfikacja jako młodocianego lambeozaura miały miejsce w 2005[2] i nie zdążyły wejść do powszechnego obiegu. W końcu Didanodon i Trachodon altidens wciąż traktuje się jako wątpliwe. Najświeższa praca przypisuje Didanodon do Lambeosaurus bez komentarza, Trachodon altidens zaś umieszcza oddzielnie w tabeli grupującej nomina dubia[3].
Paleoekologia

Lambeosaurus lambei i L. magnicristatus z formacji Dinosaur Park należały do zróżnicowanej i dobrze udokumentowanej fauny prehistorycznych zwierząt. Obejmowała ona następujące dinozaury: ceratopsy centrozaura, styrakozaura i chasmozaura, kaczodziobe prozaurolofa, grypozaura, korytozaura i parazaurolofa, tyranozauryda gorgozaura, pancerne edmontonię i euoplocefala[26]. Formację Dinosaur Park interpretuje się jako niskoreliefowe osady rzeczne i równinę zalewową, mają to być tereny bagniste o dużym wpływie morza z czasów, gdy Morze Środkowego Zachodu posuwało się na zachód[27]. Panował wtedy klimat cieplejszy, niż obecnie w Albercie, nie zdarzały się mrozy. Następowały po sobie pory suche i wilgotne. Iglaste wyraźnie dominowały wśród roślin drzewiastych. W podszycie rosły paprotniki, w tym paprocie drzewiaste, i okrytonasienne[28]. Anatomicznie podobne L. lambei, L. magnicristatus i Corythosaurus były rozdzielone czasowo, co uwidacznia się w stratygrafii. Skamieliny korytozaura znane są z niższych dwóch trzecich formacji, skamieniałości L. lambei występują w górnej jednej trzeciej, a L. magnicristatus należą do rzadkości i są obecne jedynie na szczycie, gdzie przejawia się większy wpływ morza[29].
Pożywienie

Jako hadrozaur Lambeosaurus był dużym dwu- lub czworonożnym roślinożercą. Spożywał pokarm roślinny dzięki wyspecjalizowanej czaszce zdolnej do wykonywania ruchów miażdżących analogicznych do żucia ssaków. Jego zęby podlegały ciągłej wymianie. Tworzyły baterie zębowe, z których każda zawierała ponad 100 zębów. Tylko niewielka część z nich kiedykolwiek była w użyciu. Do zrywania pokarmu zwierzę używało dzioba. Następnie trzymało go w przypominających policzki narządach. Zauropsyd pasł się na wysokości od poziomu gruntu do około 4 m[3]. Jak zauważył Robert Bakker, lambeozaurynowe dzioby były węższe od hadrozaurynowych. W związku z tym Lambeosaurus i jego krewni mogli żywić się bardziej selektywnie, niż ich szerokodziobi, bezgrzebieniaści krewni[30].
Grzebień
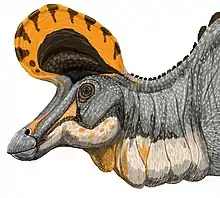
Jak inne Lambeosaurinae, w tym parazaurolof i korytozaur, Lambeosaurus miał na szczycie głowy charakterystyczny grzebień. Jego jama nosowa biegła w tył przez grzebień, w większości wydrążony. Funkcja lub funkcje grzebienia stały się przedmiotem wielu hipotez. Wedle nich mieściły się tam gruczoły solne, narząd poprawiający węch, używana w nurkowaniu rurka lub magazyn powietrza, komora rezonansowa pomagająca w wydawaniu dźwięków. Mogły też służyć odróżnianiu osobników różnych płci lub też różnych gatunków[19][31]. Funkcje społeczne, jak udział w wydawaniu dźwięków czy rozpoznawaniu się zwierząt, zdobyły sobie największą akceptację wśród wymienionych hipotez[3]. Duży rozmiar hadrozaurzych oczodołów i obecność twardówkowego pierścienia w oczach wskazują na dużą ostrość wzroku i dzienny tryb życia, w przypadku którego wzrok odgrywa dużą rolę. Słuch kaczodziobych również wydaje się dobrze rozwinięty. Istnieje co najmniej jeden przykład smukłego strzemiączka u lambeozaurynów, dotyczący blisko spokrewnionego korytozaura. Ta gadzia kosteczka słuchowa razem z dużą przestrzenią odpowiadającą błonie bębenkowej wskazuje na czułe ucho środkowe. Poza tym lagena ucha wewnętrznego hadrozaurów jest wydłużona, jak u krokodyli. Oznacza to dobrze rozwiniętą odpowiedzialną za słuch część ucha wewnętrznego[19]. Jeśli służył wydawaniu odgłosów, grzebień w dalszym ciągu mógł grać rolę w odróżnianiu się osobników różnych płci czy różnych gatunków, ponieważ zróżnicowane układy przewodów nosowych odpowiadające różnym kształtom grzebienia brałyby udział w powstawaniu różnych dźwięków[32].
Przypisy
- 1 2 3 4 5 Peter Dodson. Taxonomic implications of relative growth in lambeosaurine dinosaurs. „Systematic Zoology”. 24 (1), s. 37–54, 1975. DOI: 10.2307/2412696.
- 1 2 3 The type specimen of Tetragonosaurus erectofrons (Ornithischia: Hadrosauridae) and the identification of juvenile lambeosaurines. W: David C. Evans, Catherine F. Forster & Robert R. Reisz: Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Bloomington: Indiana University Press, 2005, s. 349–366. ISBN 0-253-34595-2.
- 1 2 3 4 5 6 7 8 9 David B. Weishampel, Jack R. Horner: Hadrosauridae. W: David B. Weishampel, Peter Dodson, Halszka Osmólska: The Dinosauria. Wyd. 1. Berkeley: University of California Press, 1990, s. 534–561. ISBN 0-520-06727-4.
- ↑ Richard Swann Lull, Nelda E. Wright: Hadrosaurian Dinosaurs of North America. Geological Society of America, 1942, s. 193–194, seria: Geological Society of America Special Paper 40.
- 1 2 3 4 5 6 David C. Evans, Robert R Reisz. Anatomy and relationships of Lambeosaurus magnicristatus, a crested hadrosaurid dinosaur (Ornithischia) from the Dinosaur Park Formation, Alberta. „Journal of Vertebrate Paleontology”. 27 (2), s. 373–393, 2007. DOI: 10.1671/0272-4634(2007)27[373:AAROLM]2.0.CO;2.
- ↑ Richard Swann Lull & Nelda E. Wright: Hadrosaurian Dinosaurs of North America. 1942, s. 227–228.
- ↑ Richard Swann Lull & Nelda E. Wright: Hadrosaurian Dinosaurs of North America. 1942, s. 112–117.
- 1 2 Lawrence M. Lambe. On Vertebrata of the mid-Cretaceous of the Northwest Territory. 2. New genera and species from the Belly River Series (mid-Cretaceous). „Contributions to Canadian Paleontology”. 3, s. 25–81, 1902.
- ↑ Lawrence M. Lambe. On a new genus and species of carnivorous dinosaur from the Belly River Formation of Alberta, with a description of the skull of Stephanosaurus marginatus from the same horizon. „Ottawa Naturalist”. 28, s. 13–20, 1914.
- ↑ William A. Parks. Corythosaurus intermedius, a new species of trachodont dinosaur. „University of Toronto Studies, Geological Series”. 15, s. 1–57, 1923.
- ↑ Richard Swann Lull & Nelda E. Wright: Hadrosaurian Dinosaurs of North America. 1942, s. 178–187.
- ↑ Henry Fairfield Osborn. Distinctive characters of the mid-Cretaceous fauna. 1. New genera and species from the Belly River Series (mid-Cretaceous). „Contributions to Canadian Paleontology”. 3, s. 1–21, 1902.
- ↑ William Diller Matthew. Canadian dinosaurs. „Natural History”. 20 (5), s. 1–162, 1920.
- ↑ William A. Parks. A new genus and two new species of trachodont dinosaurs from the Belly River Formation of Alberta. „University of Toronto Studies, Geological Series”. 31, s. 1–11, 1931.
- 1 2 Charles M. Sternberg. Hooded hadrosaurs of the Belly River Series of the Upper Cretaceous. „Canada Department of Mines Bulletin (Geological Series)”. 77 (52), s. 1–37, 1935.
- ↑ Richard Swann Lull & Nelda Wright: Hadrosaurian Dinosaurs of North America. 1942, s. 1–242.
- ↑ William A. Parks. New species of trachodont dinosaurs from the Cretaceous formations of Alberta. „University of Toronto Studies, Geological Series”. 37, s. 1–45, 1931.
- ↑ John H. Ostrom. The systematic position of Hadrosaurus (Ceratops) paucidens Marsh. „Journal of Paleontology”. 38 (1), s. 130–134, 1964.
- 1 2 3 James A. Hopson. The evolution of cranial display structures in hadrosaurian dinosaurs. „Paleobiology”. 1 (1), s. 21–43, 1975.
- 1 2 D.P. Simpson: Cassell's Latin Dictionary. Wyd. 5. London: Cassell Ltd., 1979, s. 883. ISBN 0-304-52257-0.
- ↑ John R. Horner. Upper Cretaceous dinosaurs from the Bearpaw Shale (marine) of south-central Montana with a checklist of Upper Cretaceous dinosaur remains from marine sediments in North America. „Journal of Paleontology”. 53 (3), s. 566–577, 1979.
- ↑ William J. Morris. A new species of hadrosaurian dinosaur from the Upper Cretaceous of Baja California: ?Lambeosaurus laticaudus. „Journal of Paleontology”. 55 (2), s. 453–462, 1981.
- 1 2 Terry A. Gates, Scott D. Sampson, Carlos R. Delgado de Jesús, Lindsay E. Zanno, David Eberth, René Hernandez-Rivera, Martha C. Aguillón Martínez & James I. Kirkland. Velafrons coahuilensis, a new lambeosaurine hadrosaurid (Dinosauria: Ornithopoda) from the Late Campanian Cerro del Pueblo Formation, Coahuila, Mexico. „Journal of Vertebrate Paleontology”. 27 (4), s. 917–930, 2007. DOI: 10.1671/0272-4634(2007)27[917:VCANLH]2.0.CO;2.
- ↑ Albert Prieto-Márquez, Luis M. Chiappe i Shantanu H. Joshi. The Lambeosaurine Dinosaur Magnapaulia laticaudus from the Late Cretaceous of Baja California, Northwestern Mexico. „PLoS ONE”. 7 (6), s. e38207, 2012. DOI: 10.1371/journal.pone.0038207. (ang.).
- ↑ Lambeosaurus. W: Donald F. Glut: Dinosaurs: The Encyclopedia. Jefferson, North Carolina: McFarland & Co, 1997, s. 525–533. ISBN 0-89950-917-7.
- ↑ Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loeuff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth, M.P.; & Noto, Christopher R. (2004). „Dinosaur Distribution”, in The Dinosauria (2nd), ss. 517–606.
- ↑ Eberth, David A. 2005. „The geology”, in Dinosaur Provincial Park, ss. 54–82.
- ↑ ampanian palynomorphs. W: Dennis R. Braman & Eva B. Koppelhus: Dinosaur Provincial Park. 2005, s. 101–130.
- ↑ Ornithischian Dinosaurs. W: Michael J. Ryan, David C. Evans: Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Bloomington: Indiana University Press, 2005, s. 312–348. ISBN 0-253-34595-2.
- ↑ Robert T. Bakker: The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and their Extinction. New York: William Morrow, 1986, s. 194. ISBN 0-8217-2859-8.
- ↑ Hadrosaurids II. W: David B. Norman: The Illustrated Encyclopedia of Dinosaurs: An Original and Compelling Insight into Life in the Dinosaur Kingdom. New York: Crescent Books, 1985, s. 122–127. ISBN 0-517-46890-5.
- ↑ David B. Weishampel. Acoustic analyses of potential vocalization in lambeosaurine dinosaurs (Reptilia: Ornithischia). „Paleobiology”. 7 (2), s. 252–261, 1981.

- kategoria
- klasyfikacja
- lista
- wikiprojekt
| Infrarzędy dinozaurów | |
|---|---|
| Era dinozaurów | |
| Dyscypliny powiązane |
