| Hypacrosaurus | |||
| Brown, 1913 | |||
| Okres istnienia: 70–67 mln lat temu | |||
 | |||
| Systematyka | |||
| Domena | |||
|---|---|---|---|
| Królestwo | |||
| Typ | |||
| Podtyp | |||
| Gromada | |||
| Podgromada | |||
| Nadrząd | |||
| Rząd | |||
| Podrząd | |||
| Infrarząd | |||
| (bez rangi) | iguanodony | ||
| Rodzina | |||
| Podrodzina | |||
| Plemię | |||
| Rodzaj |
hipakrozaur | ||
| Synonimy | |||
| |||
| Gatunki | |||
|
| |||
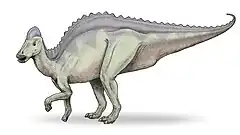
Hipakrozaur (Hypacrosaurus, "prawie najwyższy jaszczur" z greki υπο-, hypo- = mniej + ακρος, akros, wysoki, był prawie tak wysoki, jak tyranozaur[1][2] − rodzaj kaczodziobego dinozaura przypominającego wyglądem korytozaura. Łączyły je wysoki, zaokrąglony, wydrążony grzebień na głowie, u hipakrozaura nieco niższy i prostszy. Ornitopod znany jest dzięki pozostałościom dwóch gatunków żyjących pomiędzy około 76 i 68 milionami lat temu w późnokredowych Albercie w Kanadzie i Montanie w USA. Jest to ostatni kaczodzioby o wydrążonym grzebieniu, po którym pozostały dobrej jakości pozostałości w Ameryce Północnej. Pozostawał mało znany, nim w latach dziewięćdziesiątych XX wieku opisano gniazdo, jaja i wylęgające się młode gatunku H. stebingeri.
Budowa
Czaszka

Wydrążony grzebień czaszki przypomina spotykany u korytozaura, ale na szczycie jest bardziej zaostrzony, nieco niższy, szerszy w wymiarze bocznym, a w tylnej części ma niewielki kostny czubek[3]. Spłaszczony bocznie, przybiera różne kształty. U niektórych osobników gatunku typowego i u H. stebingeri wysokością nie dorównuje długości, u innych przedstawicieli H. altispinus te wymiary są sobie równe[4]. Wedle Davida Weishampela w przeciwieństwie do innych lambeozaurynów przestrzenie powietrzne w grzebieniu nie przypominały kształtem litery S (nie licząc H. altispinus)[5]. Przeciwnie – kanały te, nie łącząc się w obrębie pyska, przebiegały prosto, kończąc się wspólną komorą ułożoną pośrodkowo, łączącą się nieparzystym otworem z przestrzenią międzyoczodołową. Jednakże uczony ten użył uszkodzonych szczątków. Nowe dane uzyskane metodą tomografii komputerowej ujawniły jednak S-kształtne zagięcie, choć różniące się od spotykanego u korytozaura, w przeciwieństwie do podobnych u obu rodzajów bocznych uchyłków, nie rozciągających się poza wspomnianą pośrodkową komorą. Wedle tych badań, przeprowadzonych przez Evansa, Ridgely'ego i Witmera, dolna pętla litery S jest wydłużona, sięgając skraju czaszki. Z kolei udział w tworzeniu jego środkowej zewnętrznej powierzchni przez wyrostek kości przedszczękowej wyróżnia gatunek typowy wśród lambeozaurynów. Kość ta, duża i o złożonej budowie, od tyłu łącząca się z kością nosową, nosi pośrodkowo szew sięgający poza czubek grzebienia, jednak nie jego tylnego końca, jak u lambeozaura. Kość ta, o szerokim i łukowatym brzegu, tworzy rodzaj bezzębnego dzioba. Jej brzuszno-boczna gałąź kontaktuje się ze szczęką, kością przedoczodołową, kością łzową. Ta ostatnia to niewielka, klinowata kość usytuowana na dziobowym krańcu oczodołu, łącząca się też ze szczęka (po stronie brzusznej) i kością przedoczodołową (po grzbietowej). Z kością jarzmową tylko się styka. Korytozaur czy lambeozaur cechują się też bocznymi ciemiączkami grzebienia, których brakuje opisywanemu rodzajowi. Grzbietową i tylną część grzebienia tworzy kość nosowa. Przypada na nią od 52 do 55% powierzchni bocznej tego tworu u dorosłego zwierzęcia. Przypomina ona swój homolog u korytozaura. Wydaje się nieco zredukowana, dolna gałąź jej przedniego zakończenia jest bardzo mała lub w ogóle jej nie ma. W przedniej części nie dorównuje grubością korytozaurzej. Tam też znajduje się różniący się u poszczególnych osobników kształtem, zaostrzony lub zaokrąglony, wyrostek zachodzący bocznie na grzbietowy wyrostek kości przedszczękowej. W części dziobowej cechują ją zagłębienia, od strony brzusznej, gdzie spotyka się z kością czołową, jest poszerzona. Do autapomorfii gatunku typowego dotyczących czaszki zaliczają się m.in. otwierające się daleko na przedzie nozdrza zewnętrzne (ich kształt określa się jako łzowaty), brak pustej przestrzeni nosowej po boku grzebienia czy kąt 140° pomiędzy pyskiem i grzebieniem. Kąt ten, tworzony przez wklęsły rostralny brzeg kości przysiecznej i wznoszący się wyrostek dorsokaudalny, osiąga największą odnotowaną wartość 163° u najmniejszego z poznanych osobników. Jednak zmienia się w trakcie ontogenezy w stopniu mniejszym niż u korytozaura czy lambeozaura. Z kolei już zwężenie wspomnianych nozdrzy i otoczenie ich od tyłu przez powiększony grzbietowy brzeg kości przedszczękowej uznano za autapomorfię rodzaju Hypacrosaurus. Podobnego statusu doczekał się nierozgałęziony wyrostek łuskowy kości zaoczodołowej, pewne cechy kości jarzmowej czy brak ciemiączka pomiędzy kośćmi nosową i przedszczękową przez całe życie zwierzęcia[4].
Kość łuskowa H. altispinus jest cienka i wklęsła, o pochylonej powierzchni bocznej, jak u innych Lambeosaurinae. Ponad kością nadoczodołową spotyka swój odpowiednik po drugiej stronie czaszki. Od przodu ogranicza okno nadskroniowe, w które zagłębia się cienkim, tępym wyrostkiem swej przedniej części. Kość ta zatacza łuk w przyśrodkowo-tylnym kącie wymienionego otworu czaszki, by połączyć się z kością ciemieniową, tworzącą przyśrodkowe i przednio-przyśrodkowe ograniczenie tego okna, jak również górną część puszki mózgowej. W kierunku przednio-tylnym os parietale zajmuje mniej miejsca niż u innych kaczodziobych. Prócz obu gatunków hipakrozaura podobne występuje u Parasaurolophus walkeri. Licząca sobie 8 cm kość ta stanowi niecałe 40% odległości międzyoczodołowej, gdy u korytozaura czy lambeozaura jest to więcej niż połowa. W przeciwieństwie do poprzedniej jest to kość nieparzysta, leżąca pomiędzy kością łuskową z tyłu oraz kośćmi zaoczodołową i czołową z przodu. W tym ostatnim kierunku odchodzi od niej niewielki, różniący się rozmiarami u różnych osobników wyrostek międzyczołowy, rozwinięty lepiej od wewnętrznej strony czaszki. Zarówno kości czołowa, połączona brzusznie z homologami kości klinowej, przedczołowa, jak i zaoczodołowa nie różnią się specjalnie od spotykanych u innych korytozaurów. Tę ostatnią wyróżnia tylko kształt wyrostka ogonowego, inny nawet niż u H. stebingeri[4].

Pomiędzy tylnymi ścianami kości przedszczękowych i kością klinową podstawową leży stosunkowo duży, osiągający 52% długości szczęki lemiesz, trójkątnego kształtu, zaostrzony ku przodowi, cienki, grubszy w tylnej części, z otworem pośrodku. Cienka i trójkątna jest również kontaktująca się z nim od strony ogonowo-bocznej kość podniebienna, u gatunku typowego opisana dopiero w 2010. Na brzegu grzbietowo-ogonowym nosi panewkę dla wyrostka skrzydłowatego, przypominającego korytozaurzy, o dobrze wykształconej krawędzi brzusznej, stykający się jej przedłużeniem z ramieniem kości kwadratowej (podobnej do spotykanych u innych Lambeosaurinae), z wytworzeniem nieruchomego połączenia kości. Z kąta kości podniebiennej wystaje wyrostek dziobowo-boczny, którego brzeg brzuszny stanowi grzbietowe odgraniczenie okrągłego otworu, wedle Hornera homologicznego z oknem przedoczodołowym, zamykanego przez szczękę i ectopterygoid, wielkości dziobowego otworu szczękowego. Wspomniany wyrostek kontaktuje się z kością jarzmową. Kość jarzmowa przybiera kształt wygięty bocznie, co wiąże się z wydłużeniem czaszki, o kanciastym lub rzadziej lekko zaokrąglonym brzegu dolnym. Z tyłu od kości kwadratowej oddziela ją kość kwadratowo-jarzmowa. Wyrostek zaoczodołowy os jugale, który wraz z tylną płytką tworzy łuk wyznaczający dolne ograniczenie otworu podskroniowego, a kąt między tymi dwoma strukturami jest mniejszy niż u innych korytozaurów, przebiega w kierunku grzbietowym, by połączyć się z analogicznym wyrostkiem kości zaoczodołowej. Płytka przednia jest krótka, a jej dziobowy brzeg prosty. Dziobowy wyrotek rozpościera się w kierunku grzbietowo-brzusznym. Kształtem u H. altispinus przypomina najbardziej spotykany u Lambeosaurus magnicristatus. U H stebineri zwłaszcza u osobników młodocianych jest on szerszy i bardziej zaokrąglony niż u gatunku typowego, skraca się z wiekiem tylko nieznacznie. Przypomina w tym względzie korytozaura, L. lambei czy Parasaurolophus walkeri. Od przodu kontaktuje się z os jugale szczęka. Ta ostatnia zawiera wedle Hornera od 42 do 45, wedle Evansa zaś od 46 do 47 rodzin zębów, z których funkcjonalne zęby (po 2) znaleziono w 12. Ich ilość zależy od wielkości zwierzęcia. Kość ta cechuje się też różną liczbą różnej wielkości otworów, co stanowi cechę osobniczą[4].
Czaszka najmniejszego z odkrytych osobników (CMN 2247) mierzy około 36,6 cm długości. Największa (CMN 8675) mierzy 83,0 cm. Szczęka tego osobnika ma dokładnie taką długość, jaką czaszka CMN 2247, u którego szczęka mierzy sobie 14,9 cm[4].
Szkielet pozaczaszkowy
Długość ornitopoda szacuje się na około 9,1 m[3], masę zaś na do 4,0 tony[6]. Jak u większości kaczodziobych, jego szkielet nie wyróżnia się znacząco, aczkolwiek odróżniają go od innych pewne szczegóły budowy miednicy[7]. Prócz grzebienia hipakrozaura najłatwiej odróżnić od innych Lambeosaurinae (hadrozaurów o wydrążonych grzebieniach na głowach) dzięki wysokim wyrostkom kolczystym[3] kręgów piersiowych od 6 do 10[4]. Wspomniane wyrostki, wysokie i masywne, osiągają wysokość od 5 do 7 razy przewyższającą rozmiar zredukowanego trzonu odpowiedniego kręgu tułowia, co daje w wyniku wysoki profil pleców[3]. Należą do najwyższych wśród iguanodontów, nie licząc uranozaura.
Długą kość kulszową wraz z jej dużym, stopowato zakończonym wyrostkiem uznano nawet za jedyną pozaczaszkową autapomorfię gatunku typowego. Ostrom w 1961 opisał podobny, będący skutkiem konwergencji twór u Parasaurolophus cyrtocristatus[4]. Jak inne hadrozaury, hipakrozaur poruszał się na dwóch lub czterech nogach. Żywił się roślinami. Dwóch znanych gatunki, H. altispinus i H. stebingeri, nie można zróżnicować, przedstawiając zespół typowych dla każdego z nich autapomorfii, wobec czego H. stebingeri opisano jako formę przejściową pomiędzy wcześniejszym lambeozaurem i późniejszym hipakrozaurem[8].
Historia

Holotypowe szczątki hipakrozaura znalazł w 1910 Barnum Brown dla Amerykańskiego Muzeum Historii Naturalnej[7]. Obejmowały niekompletny szkielet pozaczaszkowy składający się z kilku kręgów i niepełnej miednicy (AMNH 5204) znad rzeki Red Deer w pobliżu Tolman Ferry[1], znaleziony po wschodniej stronie doliny rzeki Red Deer, około 6 km na północ od Tolman Bridge i 15 km na wschód od Trochu w Albercie w Kanadzie. Otaczające je skały zalicza się dziś do formacji Horseshoe Canyon powstałej w mastrychcie (kreda górna). Holotyp pochodzi prawdopodobnie z podstawy czwartej warstwy tej formacji, jego macierzysty kamieniołom datuje się na wczesny mastrycht[4]. Brown opisał rzeczone szczątki w połączeniu innymi kośćmi pozaczaszkowymi w 1913 jako nowy rodzaj, który uznał za podobny do zaurolofa[1]. Wtedy nie znano jeszcze czaszki, ale niedługo później znaleziono i opisano dwie czaszki[9].
W tamtym okresie pozostałości małych kaczodziobych o wydrążonych grzebieniach opisywano w osobnych rodzajach i gatunkach. Pierwszy z nich zapisany w historii hipakrozaura to Cheneosaurus tolmanensis bazujący na czaszce i pomieszanych kościach kończyn, kręgi i kości miednicy z formacji Horseshoe Canyon[10]. Niedługo później Richard Swann Lull i Nelda zidentyfikowali szkielet z Amerykańskiego Muzeum Historii Naturalnej (AMNH 5461) z formacji Two Medicine z Montany jako okaz Procheneosaurus[11]. Ten i inne taksony akceptowano jako pewne rodzaje przed latami siedemdziesiątymi, kiedy Peter Dodson wykazał, że z większym prawdopodobieństwem chodzi o młodociane formy innych wyróżnianych lambeozaurynów. Choć miał na myśli głównie wcześniejsze, pochodzące z formacji Dinosaur Park rodzaje Corythosaurus i Lambeosaurus, zasugerował też, jakoby Cheneosaurus obejmował pozostałości młodych osobników współczesnego mu Hypacrosaurus altispinus[12]. Pogląd ten zaakceptowano[6], ale go formalnie nie przetestowano. Pochodzący z formacji Two Medicine Procheneosaurus nie przypominał dokładnie okazów zbadanych przez Dodsona. Był znacznie bardziej podobny do nie opisanego do 1994 H. stebingeri[8].
Kolejne szczątki w formacji Horseshoe Canyon znajduje się cały czas. W 2003 odnaleziono dużą kość krzyżową wraz z długimi wyrostkami kolczystymi, która niestety uległa uszkodzeniu podczas transportu helikopterem[13].
Gatunki
H. altispinus, gatunek typowy, znany jest z 5-10 połączonych stawowo czaszek z pewnymi powiązanymi z nimi szczątkami szkieletów, od osobników młodocianych do dorosłych, H. stebingeri zaś z nieznanej dokładnie, lecz znacznej liczby osobników od zarodków po dorosłe[6]. H. altispinus uznawano szeroko za bezpośredniego potomka korytozaura[4]. Monofiletyzm grupy zawierającej tylko dwa gatunki hipakrozaura bywa podważany choćby przez pracę Suzuki et al., powtórny opis nipponozaura z 2004. Ta analiza filogenetyczna sytuuje Nipposaurus bliżej H. altispinus niż gatunek H. stebingeri[14]. Również publikacja Prieto-Marquez et al. z 2006 sugeruje parafiletyzm rodzaju[4]. Jednak Evans i Reisz w 2007 odrzucili ten pogląd[15], Evans utrzymał ten pogląd w 2010[4].
Nowym gatunkiem Hypacrosaurus stebingeri nazwano rozmaite szczątki, w tym wykluwające się młode razem z jajami i gniazdami, znalezione w szczytowej części późnokampańskiej (kreda górna) formacji Two Medicine Formation w Glacier Creek w Montanie (Stany Zjednoczone) i na granicy Alberty[8] (szczątki z południa Alberty należą do formacji Oldman[4]). Określono je jako największą kolekcję niemowlęcych szkieletów wśród wszystkich poznanych do tej pory gatunków kaczodziobych[8]. Badania tego gatunku podważyły wspomnianą wyżej tezę o tak bliskim pokrewieństwie hipakrozaura i korytozaura. Wedle pracy Hornera z 1992 H. stebingeri jako gatunek "przejściowy" leży na swojej linii ewolucyjnej pomiędzy bardziej bazalnym lambeozaurem i kończącym tą gałąź H. altispinus (anageneza)[4].
Systematyka
Fragment kladogramu autorstwa Evans & Reisz (2070)[4], prezentujący drobne różnice w zależności od wykorzystanej metody:
Inne wyniki zaprezentowali w 2011 Sullivan et al.[16] (jedno z 4 rozważanych drzew, bazujące na kości ramiennej i jarzmowej), zmodyfikowano:
|
Hypacrosaurus należał do dinozaurów kaczodziobych z grupy lambeozaurów. Sklasyfikowano go tak na podstawie opisu jego czaszki[9]. W obrębie Lambeosaurinae jego najbliżsi krewni to Lambeosaurus i Corythosaurus[6]. Jack Horner i Philip J. Currie zasugerowali w 1994, że H. stebingeri stanowi formę przejściową pomiędzy Lambeosaurus i H. altispinus[8]. Oznaczałoby to ewolucję poprzez anagenezę. Zjawisko to uznawano niekiedy za dominujący typ ewolucji późkokredowych dinozaurów[4]. W 1989 Michael K. Brett-Surman zaproponował uznanie Hypacrosaurus i Corythosaurus za ten sam rodzaj[17]. Rodzaje te, zwłaszcza Corythosaurus i Hypacrosaurus, uznaje się za gałąź lambeozaurynów charakteryzującą się obecnością hełmów na głowach. Choć redeskrypcja Nipponosaurus autorstwa Suzuki et al. z 2004 doszukała się bliskiego pokrewieństwa pomiędzy Nipponosaurus i Hypacrosaurus stebingeri i parafiletyczności rodzaju Hypacrosaurus[14], pogląd ten odrzucono w późniejszej, bardziej wyczerpującej powtórnej analizie Lambeosaurinae, wedle której oba gatunki hipakrozaura tworzą klad, do którego nie należy Nipponosaurus, a Corythosaurus i Olorotitan to ich najbliżsi krewni[15]. W 1990 Chapman i Brett Surman ukulu pojęcie nieformalnych kladów korytozaura i parazaurolofa. Ten pierwszy obejmował korytozaura, hipakrozaura, lambeozaura i nipponozaura. Oba pojęcia zmodyfikował Godefroit w 2004. W kladzie korytozaura umieścił trzy wymienione wcześniej rodzaje, dodając opisanego w 2003 olorotytana. Nie wymienił tu nipponozaura. W 2007 Evans i Reisz sformalizowali go, nadając mu nazwę Corythosaurini. Jednakże Sullivan i współpracownicy w 2011, powołując się na Międzynarodowy Kodeks Nomenklatury Zoologicznej, którego artykuł 37.1 stanowi, że w przypadku podziału rodziny nazwy taksonów podrzędnych zawierających rodzaj typowy powinno tworzyć się od nazwy tego rodzaju, uznają za prawidłową nazwę Lambeosaurini, nie Corythosaurini. W plemieniu tym umieszczają cztery rodzaje kladu korytozaura wedle Godefroita i dodatkowo Velafrons[16].
Paleoekologia
Obfite pozostałości Hypacrosaurus altispinus[18] pochodzą z formacji Horseshoe Canyon, gdzie stanowią w niektórych miejscach drugie najpospolitsze skamieliny[19][13]. W odróżnieniu od skrajnych pierwszej i piątej środkowe trzy podjednostki reprezentują klimat chłodny i suchy[13]. To właśnie one odpowiadają obszarowi w dolinie rzeki Red Deer, gdzie odnaleziono pozostałości hipakrozaura. Szczątki tego zwierzęcia datuje się na okres od 70,44 ± 0,17 do 67 milionów lat temu[4]. Ogólnie w skałach tej formacji znaleziono sześć ornitopodów: prócz hipakrozaura[13] zaurolofa[19] (Saurolophus osborni[18]) Thescelosaurus neglectus, Parksosaurus warreni oraz dwa gatunki edmontozaura (Edmontosaurus regalis Lambe, 1917 i E. annectens (Marsh, 1892)[18]). Prócz nich w skałach Horseshoe Canyon znaleziono też pozostałości, pachycefalozaury Stegoceras edmontonense i S. validum, dinozaury rogate Montanoceratops cerorhynchus, Anchiceratops ornatus, Arrhinoceratops brachyops, Pachyrhinosaurus canadensis i pewne niezidentyfikowane szczątki ceratopsów, jak również ich ślady, ankylozaury Euoplocephalus tutus, Edmontonia longiceps i Panoplosaurus sp[20] (praca Larsona wymienia też niesklasyfikowane dokładniej szczątki z rodziny Ankylosauridae[13]). Z teropodów znaleziono tam celurozaura incertae sedis Ricardoestesia gilmorei, owiraptorozaura Chirostenotes pergracilis i Caenagnathus sp., ornitomimozaury Ornithomimus edmontonensis, Struthiomimus altus[20] i Dromiceiomimus[13], jak również niezidentyfikowane bliżej Ornithomimidae[20], alwarezaura Albertonykus borealis[13], przedstawicieli troodontów[20], jak Troodon czy Paronychodon[13], tyranozauroidy Aublysodon sp., Albertosaurus sarcophagus, Daspletosaurus sp. i nieokreślone bliżej pozostałości, dromeozaury, jak nieopisany jeszcze gatunek dromeozaura, Saurornitholestes[20], Atrociraptor marshalli[13] i inne, niezidentyfikowane bądź nieopisane, prócz tego członków nadrodziny Avialae[20]. Prócz dinozaurów odnaleziono tam pozostałości wielu innych stworzeń. Wśród łuskonośnych wymienia się Leptochamops z Borioteiioidea (Scincomorpha) i niezidentyfikowanych przedstawicieli tego kladu, z champsozaurów Champsosaurus. Żółwie reprezentowali przedstawiciele skorpuchowatych. Żyły tam płazy z grupy skaczących: Palaeobatrachus occidentalis oraz pewne niezidentyfikowane bliżej Salientia oraz płazy ogoniaste: Scapherpetontidae, zarówno niesklasyfikowane dokładniej, jak i rodzaj Scapherpeton, Opisthotriton z rodziny Batrachosauroididae. Ryby kostnoszkieletowe z tej formacji to niezidentyfikowani bliżej przedstawiciele kladu Acanthomorpha, rzędu elopsokształtnych, rodzin Sorbinichtyidae, szczupakowatych, jesiotrowatych i wiosłonosowatych, a także Cyclurus, Belonostomus, Coriops, Oldmanesox, Estesesox. Wśród ryb chrzęstnoszkieletowych znaleziono Hybodus, Cretoxyrhina, Ischyrhiza, Myledaphus. Typ mięczakow reprezentuje Viviparus prudentius[13].

Dinozaury tej formacji określa się czasami terminem Edmontonian odnoszącym się zwykle do lądowej fauny ssaków[21]. Różnią się one od Dinosauria z formacji wyższych i niższych[22]. Formacja Horseshoe Canyon wykazuje znaczny wpływ morza z powodu przemieszczania się (transgresji) Morza Środkowego Zachodu[21], płytkiego morza rozpościerającego się pośrodku Ameryki Północnej przez większość okresu kredowego[22]. H. altispinus mógł preferować tereny bardziej lądowe[6].
Nieco starszą formację Two Medicine, macierzystą dla H. stebingeri, zamieszkiwały także dobrze znany gniazdujący hadrozaur majazaura i inny kaczodzioby prozaurolof, hipsylofodon orodrom, ceratopsy achelozaur, einiozaur, brachyceratops i Styracosaurus ovatus, ankylozaury edmontonia i euoplocefal, troodon troodon, tyranozaur daspletozaur, cenagnat chirostenot, dromeozaury bambiraptor i zaurornitolest[20]. Tereny te leżały nieco dalej od Morza Środkowego Zachodu. Panował tam klimat w większym stopniu lądowy, bardziej gorący i suchy[23].
Do H. stebingeri przypisuje się też szczątki z formacji Oldman na południu Alberty[4].
Paleobiologia
Jako dinozaur kaczodzioby, hipakrozaur był poruszającym się na dwu lub czterech łapach roślinożercą żywiącym się różnorodnymi roślinami. Budowa jego czaszki umożliwiała miażdżące ruchy przypominające żucie. Zęby, upakowane setkami w baterie zębowe, używane były tylko w niewielkiej części i stale zastępowane przez nowe. Ptasiomiedniczny zrywał pokarm swym szerokim dziobem, po czym kierował go do narządów przypominających policzki. Zwierzę pasło się na roślinach o przedziale wysokości sięgającym od poziomu gruntu do 4 m[6].
Gniazda i wzrost

H. stebingeri składał jaja kształtu zbliżonego do kulistego, mierzące 20 na 18,5 cm. Badania Ericksona i współpracowników (2017) nad skamieniałymi zarodkami H. stebingeri sugerują, że okres inkubacji jaj u tego dinozaura wynosił co najmniej 171,47 dni[24]. Znalezione zarodki miały długość 60 cm, wylęgając się osiągały już około 1,7 m długości. Zarodki i młode cechowały się głębokimi czaszkami o nieznacznym tylko rozwoju kości mających utworzyć w przyszłości grzebienie[8]. W następnych latach największy względny wzrost dotyczył kości nosowej i wyrostków kości przedszczękowej. Wyrostek tylno-boczny, kształtu lancetowatego lub ćwiartki koła, sytuuje się przed wyrostkiem tylno-grzbietowym, zasłanianym częściowo przez kość nosową, w przeciwieństwie do lambeozaura nie rozdziela jednak tych kości. Połączenie stawowe pomiędzy wspomnianymi kośćmi, początkowo leżące przed oczodołem, zmieniało pozycję, by znaleźć się nad nim. Z biegiem czasu zmieniała się ilość kości tworzących puszkę mózgową, gdyż kości łączyły się ze sobą. Przykład stanowi tutaj zrośnięcie kości przedklinowej i oczodołowo-klinowej (tworzą one dziobową część bocznej ściany mózgoczaszki, przez tę drugą przebiegają nerwy wzrokowy, okoruchowy i bloczkowy), które nie zaszło jeszcze u osobnika oznaczonego jako CMN 2247[4]. Wzrost przebiegał szybciej niż w przypadku aligatorów, porównywalnie z bezgrzebieniowcami, przez kilka lat, o czym wiadomo na podstawie obserwacji ilości tkanki kostnej pomiędzy pierścieniami zatrzymanego wzrostu na kościach (analogicznie do słojów drzewnych)[25]. Wedle badań Lisy Cooper i współpracowników H. stebingeri osiągał dojrzałość płciową w wieku dwóch-trzech lat, osiągając swe pełne rozmiary w wieku dziesięciu-dwunastu lat. Obwód kości udowej w postulowanym czasie osiągnięcia dojrzałości płciowej stanowił tylko 40% tego obwodu u zwierząt w pełni wyrośniętych. Postulowane tempo wzrostu H. stebingeri wyprzedza spotykane u tyranozaurów, żywiących się hipakrozaurem, jak albertozaur czy tyranozaur. Szybko rosnący kaczodzioby miał większe szanse osiągnąć chroniące go rozmiary, a wczesne rozpoczęcie rozrodu stanowiło dodatkową korzyść dla zwierzęcia stanowiącego zdobycz[26].
Grzebień
Wydrążony grzebień hipakrozaura pełnił najprawdopodobniej funkcje społeczne: mógł stanowić wizualny sygnał pozwalający zwierzętom na rozpoznanie gatunku i płci innych osobników, a także stanowić komorę rezonansową pomocną w wydawaniu dźwięków[6]. Grzebień i związane z nim komory powietrzne odegrały także rolę w debacie na temat stałocieplności dinozaurów, zwłaszcza tyczących się małżowin nosowych. Są to cienkie kości lub struktury chrzęstne dwóch typów, odgrywające dwie role. Małżowiny węchowe spotyka się także u współczesnych czworonogów, biorą one udział w odczuwaniu zapachów. Małżowiny oddechowe chronią zwierzę przed utratą wody i występują jedynie u ptaków i ssaków, współczesnych kręgowców stałocieplnych i ciepłokrwistych. Z uwagi na znacznie szybszy metabolizm kręgowce te oddychają znacznie częściej niż stworzenia ektodermiczne tej samej wielkości, co wiąże się ze znacznie większą stratą wody[27]. Ruben i inni w 1996 dzięki tomografii komputerowej doszli do wniosku, że małżowin takich nie było u nanotyrana, ornitomima i hipakrozaura. Nie stanowią więc dowodu na ciepłokrwistość tych zwierząt[28]. Nie świadczą jednak o jej braku, gdyż u niektórych stałocieplnych małżowiny uległy redukcji lub zanikły[27].
Przypisy
- 1 2 3 Barnum Brown. A new trachodont dinosaur, Hypacrosaurus, from the Edmonton Cretaceous of Alberta. „Bulletin of the American Museum of Natural History”. 32 (20), s. 395–406, 1913. [dostęp 2007-05-02].
- ↑ Deciphering duckbills. W: Benjamin S. Creisler: Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs. Bloomington and Indianapolis: Indiana University Press, 2007, s. 185–210. ISBN 0-253-34817-X.
- 1 2 3 4 Richard Swann Lull, Nelda E. Wright: Hadrosaurian Dinosaurs of North America. Geological Society of America, 1942, s. 206–208, seria: Geological Society of America Special Paper 40.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 David C. Evans. Cranial anatomy and systematics of Hypacrosaurus altispinus, and a comparative analysis of skull growth in lambeosaurine hadrosaurids (Dinosauria: Ornithischia). „Zoological Journal of the Linnean Society”. 159, s. 398-434, 2010-06-26. The Linnean Society of London. DOI: 10.1111/j.1096-3642.2009.00611.x. (ang.).
- ↑ David B. Weishampel. The nasal cavity of lambeosaurine hadrosaurids (Reptilia:Ornithischia): comparative anatomy and homologies. „Journal of Paleontology”. 55 (5), s. 1046–1057, 1981.
- 1 2 3 4 5 6 7 David B. Weishampel, Jack R. Horner: Hadrosauridae. W: David B. Weishampel, Peter Dodson, Halszka Osmólska: The Dinosauria. Wyd. 1. Berkeley: University of California Press, 1990, s. 534–561. ISBN 0-520-06727-4.
- 1 2 Hypacrosaurus. W: Donald F. Glut: Dinosaurs: The Encyclopedia. Jefferson, North Carolina: McFarland & Co, 1997, s. 478–482. ISBN 0-89950-917-7.
- 1 2 3 4 5 6 Embryonic and neonatal morphology and ontogeny of a new species of Hypacrosaurus (Ornithischia, Lambeosauridae) from Montana and Alberta. W: John R. Horner, Phillip J. Currie: Dinosaur Eggs and Babies. Cambridge: Cambridge University Press, 1994, s. 312–336. ISBN 0-521-56723-8.
- 1 2 Charles Whitney Gilmore. On the genus Stephanosaurus, with a description of the type specimen of Lambeosaurus lambei, Parks. „Canada Department of Mines Geological Survey Bulletin (Geological Series)”. 38 (43), s. 29–48, 1924.
- ↑ Lawrence M. Lambe. On Cheneosaurus tolmanensis, a new genus and species of trachodont dinosaur from the Edmonton Cretaceous of Alberta. „The Ottawa Naturalist”. 30 (10), s. 117–123, 1917.
- ↑ William Diller Matthew. Canadian dinosaurs. „Natural History”. 20 (5), s. 1–162, 1920.
- ↑ Peter Dodson. Taxonomic implications of relative growth in lambeosaurine dinosaurs. „Systematic Zoology”. 24 (1), s. 37–54, 1975. DOI: 10.2307/2412696.
- 1 2 3 4 5 6 7 8 9 10 Derek W. Larson, Donald B. Brinkman, Phil R. Bell. Faunal assemblages from the upper Horseshoe Canyon Formation, an early Maastrichtian cool-climate assemblage from Alberta, with special reference to the Albertosaurus sarcophagus bonebed. „Canadian Journal of Earth Sciences”. 47, s. 1159-1181, 2010. DOI: 10.1139/E10-005. (ang.).
- 1 2 Daisuke Suzuki, David B. Weishampel, Nachio Minoura. Nipponosaurus sachaliensis (Dinosauria; Ornithopoda): anatomy and systematic position within Hadrosauridae. „Journal of Vertebrate Paleontology”. 24 (1), s. 145–164, 2004. DOI: 10.1671/A1034-11.
- 1 2 David C. Evans, Robert R. Reisz. Anatomy and relationships of Lambeosaurus magnicristatus, a crested hadrosaurid dinosaur (Ornithischia) from the Dinosaur Park Formation, Alberta. „Journal of Vertebrate Paleontology”. 27 (2), s. 373–393, 2007. DOI: 10.1671/0272-4634(2007)27[373:AAROLM]2.0.CO;2.
- 1 2 Robert M. Sullivan, Steven E. Jasinski, Merrilee Guenther & Spencer G. Lucas. The First Lambeosaurin (Dinosauria, Hadrosauridae, Lambesaurinae) from the Upper Cretaceous Alamo Formation (Naashoibito Member), San Juan Basin, New Mexico. „Fossil Record 3. New Mexico Museum of Natural History and Science”. 53, s. 405-417, 2011. (ang.).
- ↑ Michael K. Brett-Surman: A revision of the Hadrosauridae (Reptilia:Ornithischia) and their evolution during the Campanian and Maastrichtian. Ph.D. dissertation. Washington, D.C.: Graduate School of Arts and Sciences of The George Washington University, 1989, s. 1–272.
- 1 2 3 Tamaki Sato & Xiao-Chun Wu. Review of Plesiosaurians (Reptilia: Sauropterygia) from the Upper Cretaceous Horseshoe Canyon Formation in Alberta, Canada. „Paludicola”. 5 (4), s. 150–169, 2006-06. Rochester Institute of Vertebrate Paleontology. (ang.).
- 1 2 Robert M. Sullivan, Spencer G. Lucas. The Kirtlandian land-vertebrate "age"–faunal composition, temporal position and biostratigraphic correlation in the nonmarine Upper Cretaceous of western North America. „Late Cretaceous vertebrates from the Western Interior. New Mexico Museum of Natural History and Science Bulletin”. 35, s. 7-30, 2006. (ang.).
- 1 2 3 4 5 6 7 David B. Weishampel, Paul M. Barrett, Rodolfo A. Coria, Jean Le Loueff, Xu Xing, Zhao Xijin; Ashok Sahni, Elizabeth M.P. Gomani & Christopher N. Noto: Dinosaur distribution. W: David B. Weishampel, Peter Dodson, Halszka Osmólska: The Dinosauria. Wyd. 1. Berkeley: University of California Press, 1990, s. 517–606. ISBN 0-520-06727-4.
- 1 2 Peter Dodson: The Horned Dinosaurs: A Natural History. Princeton: Princeton University Press, 1996, s. 14–15. ISBN 0-691-05900-4.
- 1 2 Peter Dodson: The Horned Dinosaurs: A Natural History. Princeton: Princeton University Press, 1996, s. 14–15. ISBN 0-691-05900-4.
- ↑ Raymond R. Rogers. Taphonomy of three dinosaur bone beds in the Upper Cretaceous Two Medicine Formation of northwestern Montana: evidence for drought-related mortality. „PALAIOS”. 5 (5), s. 394–413, 1990. DOI: 10.2307/3514834.
- ↑ Gregory M. Erickson, Darla K. Zelenitsky, David Ian Kay i Mark A. Norell. Dinosaur incubation periods directly determined from growth-line counts in embryonic teeth show reptilian-grade development. „Proceedings of the National Academy of Sciences of the United States of America”. 114 (3), s. 540–545, 2017. DOI: 10.1073/pnas.1613716114. (ang.).
- ↑ Lisa N. Cooper, John R. Horner. Growth rate of Hypacrosaurus stebingeri as hypothesized from lines of arrested growth and whole femur circumference. „Journal of Vertebrate Paleontology”. 19 (3, Suppl.), s. 35A, 1999.
- ↑ Lisa N. Cooper, Andrew H. Lee, Mark L. Taper & John R. Horner. Relative growth rates of predator and prey dinosaurs reflect effects of predation. „Proceedings of the Royal Society B”. 275 (1651), s. 2609–2615, 2008. DOI: 10.1098/rspb.2008.0912. PMID: 18682367.
- 1 2 Chinsamy, Anusuya & Hillenius, Willem J. (2004). "Physiology of nonavian dinosaurs". The Dinosauria, wydanie 2. 643-659.
- ↑ J. A. Ruben, Hillenius, W.J.; Geist, N.R.; Leitch, A.; Jones, T.D.; Currie, P.J.; Horner, J.R.; & Espe, G., III. The metabolic status of some Late Cretaceous dinosaurs. „Science”. 273 (5279), s. 1204–1207, 08 1996. DOI: 10.1126/science.273.5279.1204.

- kategoria
- klasyfikacja
- lista
- wikiprojekt
| Infrarzędy dinozaurów | |
|---|---|
| Era dinozaurów | |
| Dyscypliny powiązane |
