Ewolucja układu nerwowego – powstanie i rozwój układu nerwowego na drodze zmian ewolucyjnych u zwierząt. Neurony rozwinęły się jako wyspecjalizowane komórki przesyłające sygnały elektryczne u zwierząt wielokomórkowych, adaptując mechanizm potencjału czynnościowego, obecnego już u ruchliwych jednokomórkowych czy kolonialnych eukariotów. Najpierw powstała prosta sieć komórek nerwowych spotykana u zwierząt takich jak parzydełkowce. Następnie u zwierząt o symetrii dwubocznej pojawiły się pnie nerwowe: pień nerwowy brzuszny u większości bezkręgowców oraz pień nerwowy grzbietowy, otoczony struną grzbietową u strunowców. Bilateralizacja doprowadziła do ewolucji mózgu w procesie zwanym cefalizacją.
Zrekonstruowanie ewolucji układu nerwowego nie należy do zadań łatwych. Nauka poznała nieliczne tylko formy zamieszkujące Ziemię w przeszłości, a dostępny nauce zapis kopalny często nie zachowuje informacji o układzie nerwowym – jego ślad zachowuje się tylko wyjątkowo. W efekcie rozwój tego układu opisuje się na podstawie anatomii porównawczej i embriologii, posiłkując się prawem biogenetycznym. Badając rozwój układu nerwowego u zwierząt dziś występujących i odnosząc jego budowę u najbardziej podobnych zwierząt dzisiejszych, wnioskuje się o jego budowie u zwierząt wymarłych i w ten sposób odtwarza jego rozwój[1].
Prekursorzy
Potencjał czynnościowy, niezbędny dla aktywności neuronalnej, wyewoluował już u jednokomórkowych eukariotów. Wykorzystują one raczej potencjał wynikający z nierównowagi stężeń kationów wapnia, niż sodu, ale ten mechanizm został prawdopodobnie zaadaptowany do przekazywania sygnałów elektrycznych przez neurony zwierząt wielokomórkowych. W pewnych koloniach eukariontów, jak Obelia, sygnały elektryczne ulegają propagacji nie tylko przez sieci nerwowe, ale także przez komórki nabłonkowe wspólnego układu pokarmowego kolonii[2].
Gąbki
W środkowej warstwie ciała gąbek znajdują się wielobiegunowe komórki o długich, cienkich wypustkach. Komórki łączą się ze sobą za pomocą tych wypustek[3]. Gąbki nie wytwarzają jednak komórek łączących się między sobą za pomocą synaps. Nie mając neuronów, nie mają też układu nerwowego. Dysponują jednak pewnymi homologami licznych genów grających kluczowe role w funkcji synaps. Niedawne badania wykazały, że w komórkach gąbek ulega ekspresji grupa białek gromadzących się razem i tworzących w ten sposób strukturę przypominającą błonę postsynaptyczną, odbierającą sygnały przekazywane przez synapsę[4]. Jednak funkcja tej struktury pozostaje niejasna. Choć komórki gąbek nie wykazują transmisji synaptycznej, komunikują się z innymi poprzez fale wapniowe i inne impulsy, które mediują proste czynności, jak skurcz całego ciała[5].
Sieć nerwowa
Parzydełkowce, żebropławy i spokrewnione z nimi zwierzęta wytwarzają rozsiane sieci nerwowe, bez ośrodkowego układu nerwowego[6]. Jednakże u tych zwierząt można mówić już o układzie nerwowym[1]. U większości meduz sieć nerwowa rozmieszczona jest mniej lub bardziej równomiernie w całym ciele. U żebropławów koncentruje się w okolicy otworu gębowego. Sieć nerwowa składa się z neuronów czuciowych odbierających bodźce chemiczne, dotykowe, wzrokowe, motoneuronów wywołujących skurcze ściany ciała oraz interneuronów wykrywających wzorce aktywności neuronów czuciowych i przesyłających sygnały do grup motoneuronów[6]. Wypustki czuciowe jamochłonów rozmieszczone są w wewnętrznej i zewnętrznej warstwie ciała. Pojawiają się też proste narządy zmysłów: wzroku i równowagi (statocysty). W ektodermie istnieją również komórki nabłonkowo-zmysłowe. Z kolei wypustki nerwowe łączą się z komórkami mięśniowymi[1]. W niektórych przypadkach grupy interneuronów gromadzą się w odrębne zwoje[6].
U promienistych układ nerwowy rozwinął się w sposób względnie niezorganizowany. W przeciwieństwie do zwierząt dwubocznie symetrycznych te o symetrii promienistej cechują się tylko dwoma pierwotnymi warstwami komórek: ektodermą i endodermą. Neurony tworzą się ze specjalnej grupy ektodermalnych komórek prekursorowych, z których powstaje także każdy inny rodzaj komórek ektodermalnych[7].
Pnie nerwowe

Znacząca większość istniejących obecnie zwierząt ma symetrię dwuboczną. Ich lewa i prawa strona ciała są mniej więcej swymi odbiciami lustrzanymi. Uważa się, że wszystkie zwierzęta o symetrii dwubocznej pochodzą od wspólnego przodka o robakowatym ciele, który pojawił się w okresie ediakaru, 550–600 milionów lat temu[8]. Zasadniczo ciało takiego zwierzęcia jest rurą z wydrążonym jelitem przebiegającym od otworu gębowego do odbytowego i pniem nerwowym z poszerzeniem (zwojem) dla każdego segmentu, szczególnie dużym na przedzie, nazywanym mózgiem.

Nawet ssaki, w tym człowiek rozumny, zachowują segmentowaną, dwubocznie symetryczną budowę na poziomie układu nerwowego. Rdzeń kręgowy wiąże się z szeregiem segmentalnych zwojów, od których odchodzą nerwy ruchowe i czuciowe zaopatrujące właściwe im dermatomy i leżące wewnątrz mięśnie. W przypadku kończyn plan unerwienia jest złożony, jednak na tułowiu zachowań jest wzór szeregu wąskich pasów. 3 szczytowe segmenty należą do mózgu, tworzą się z nich przodomózgowie, śródmózgowie i tyłomózgowie[9].
Zwierzęta dwubocznie symetrycznie dzieli się, bazując na ich rozwoju embrionalnym, na 2 duże grupy o randze wyższej od typów: pierwouste i wtórouste[10]. Te ostatnie obejmują takie bezkręgowce, jak szkarłupnie, półstrunowce i Xenoturbellida[11]. Pierwouste, grupa bardziej zróżnicowana, obejmują m.in. stawonogi, mięczaki i różnorodne typy zwierząt o robakowatym ciele. Pomiędzy tymi dwoma grupami występują ważna różnica w miejscu zajmowanym przez układ nerwowy w obrębie ciała. Pierwouste mają pień nerwowy na brzusznej (zazwyczaj dolnej) stronie ciała, podczas gdy u wtóroustych leży on po grzbietowej (zazwyczaj górnej) stronie ciała. W rzeczywistości liczne cechy ciał są u tych grup wzajemnie odwrotne, w tym wzorce ekspresji pewnych genów wykazujące gradient grzbietowo-brzuszny. Większość anatomów obecnie uważa ciała przedstawicieli tych dwóch grup za wzajemnie odwrócone. Hipotezę tą zaproponował jako pierwszy Geoffroy Saint-Hilaire dla owadów, porównując je z kręgowcami. Dlatego owady mają pnie nerwowe w brzusznej części ciała, podczas gdy u wszystkich kręgowców rdzeń kręgowy ulokowany jest w części grzbietowej, w obu przypadkach pośrodkowo[12].
Robaki
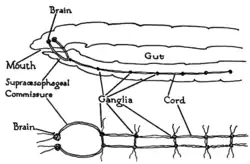
Najprostsze zwierzęta dwubocznie symetrycznie to robaki. Prezentują one podstawową strukturę układu nerwowego o symetrii dwubocznej w najprostszy sposób.
Ośrodkowy układ nerwowy pojawia się po raz pierwszy u robaków płaskich. Występuje u nich parzysty zwój nerwowy, od którego odchodzą czuciowe i ruchowe włókna nerwowe. U zwierząt tych komórki nerwowe obecne są także w nerwach[13].
Bardziej zorganizowany układ nerwowy prezentują również określane niegdyś mianem robaków pierścienice[13]. Dżdżownica ma podwójny pień nerwowy brzuszny biegnący przez całą długość jej ciała, od jamy ustnej do końca ciała. Pnie nerwowe łączą się z biegnącymi poprzecznie nerwami, przypominającymi szczeble w drabinie[14], widać tutaj przykład metamerii[13]. Biorą one udział w koordynacji obu stron ciała zwierzęcia. Dwa zwoje w okolicy końca głowy pełnią funkcję prostego mózgu[14]. W obrębie tych zwojów perikariony znajdują się od zewnątrz, środek wypełnia neuropil[13]. Fotoreceptory na plamkach ocznych dostarczają informacji o świetle bądź ciemności[14]. U wyższych pierścienic oba zwoje nerwowe są połączone, tworzy się nieparzysty brzuszny łańcuch nerwowy[13].
Układ nerwowy bardzo małych rozmiarów nicienia Caenorhabditis elegans został zmapowany aż do poziomu synaps. Uwzględniono każdy neuron i jego linię komórkową, poznano większość, jeśli nie wszystkie połączenia neuronalne. U tego gatunku układ nerwowy wykazuje dymorfizm płciowy. Występują dwie płcie: samce i hermafrodyty, różniące się ilością neuronów i gruami komórek nerwowych obsługujących czynności specyficzne dla płci. C. elegans samce mają dokładnie 383 neurony, a hermafrodyty dokładnie 302[15].
Stawonogi

Stawonogi, jak owady czy pajęczaki, posiadają układ nerwowy zbudowany z serii zwojów połączonych brzusznym pniem nerwowym, tworzonym przez 2 połączenia biegnące przez całą długość brzusznej strony ciała[16]. Mózg, najlepiej rozwinięty u owadów społecznych, jak mrówki, pszczoły, termity[13], leży w segmencie głowowym, mówi się też o zwoju nadprzełykowym. Dzieli się na protocerebrum, deutocerebrum i tritocerebrum. Zaraz za mózgiem znajduje się zwój podprzełykowy, zbudowany z trzech par połączonych ze sobą zwojów. Kontroluje on aparat gębowy, gruczoły ślinowe i pewne mięśnie. Wiele stawonogów posiada dobrze rozwinięte narządy zmysłów, w tym stanowiące zmysł wzroku oczy złożone oraz czułki zapewniające węch i wykrywające feromony. Informacje odbierane przez te narządy zmysłów trafiają do mózgu.
Podwójne zwoje metameryczne mogą łączyć się w pojedynczy zwój[13].
U owadów wiele neuronów posiada ciała rozmieszczone na obrzeżach mózgu, pasywne elektrycznie. Rola perikarionów polega jedynie na zapewnianiu wsparcia metabolicznego, nie bierze zaś udziału w przekazywaniu sygnałów. Włókna ptotoplazmatyczne biegną przez ciało komórki nerwowej i rozgałęziają się obficie, pewne ich części przekazują sygnały, a inne je zbierają. Wobec tego większa część owadziego mózgu zbudowana jest z pasywnych perikarionów zgrupowanych na obwodzie, podczas gdy przesyłanie sygnałów odbywa się w plątaninie włókien protoplazmatycznych (neuropile) ulokowanej w centrum[17].
Cewa nerwowa strunowców
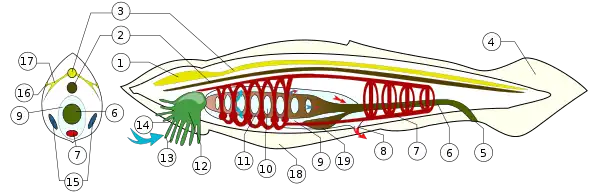
1) pęcherzyk mózgowy 2) struna grzbietowa 3) cewa nerwowa 16) komórka światłoczuła 17) nerwy
Pośród wtóroustych prócz szkarłupni o symetrii promienistych znajdują się strunowce. Zwierzęta te wykształciły strunę grzbietową, a ich układ nerwowy tworzą cewę nerwową pochodzenia ektodermalnego. U osłonic wymienione dwa narządy istnieją jedynie na początku ontogenezy, w dalszym jej ciągu zanikają (wyjątek stanowią appendikularie, u których zachowuje się strun grzbietowa, a cewa nerwowa przekształca się w pień nerwowy grzbietowy, budowany przez zwoje nerwowe połączone włóknami nerwowymi)[13].
U należącego do bezczaszkowców lancetnika w czasie jego rozwoju embrionalnego z leżącej grzbietowo w stosunku do struny grzbietowej ektodermy tworzącej płytę nerwową powstaje najpierw rynienka nerwowa, która daje początek cewie nerwowej. Zarówno struna grzbietowa, jak i cewa nerwowa nie zanikają przez całe życie zwierzęcia, co więcej, cewa nerwowa tworzy rdzeń kręgowy, porównywany do podłużnego pręta, trójkątnego w przekroju (z uchyłkiem po stronie grzbietowej w efekcie niepełnego zrośnięcie się brzegów rynienki), wydrążonego w środku. Podłużnny kanał w jego wnętrzu wyściełają komórki ependymalne. Z rdzenia odchodzą metameryczne korzenie brzuszne, naprzemiennie z korzeniami grzbietowymi. Pod względem histologicznym uwagę zwracają komórki Rhodego, należące do komórek kojarzeniowych (umożliwiają komunikację pomiędzy przednimi a tylnymi komórkami ruchowymi)[18].
U lancetnika pojawia się też pęcherzyk czołowy, odpowiednik pęcherzyka mózgowego kręgowców. Wyróżnia go histologia: tworzy go jednowarstwowy nabłonek migawkowy. Narządy zmysłu wzroku stanowią oczka, czyli pojedyncze komórki swiatłoczułe, leżące w rdzeniu. Uważa się jednak, że brak oczu pojawił się u lancetnika wtórnie, jako że zwierzę to większość swego życia spędza zagrzebane w piasku dna morskiego[19].
U kręgowców już w trakcie rozwoju zarodkowego z proksymalnej części cewy nerwowej powstają 3 pęcherzyki. Rozwijają się z nich przodomózgowie, śródmózgowie i tyłomózgowie. Przodomózgowie dzieli się dalej, tworząc kresomózgowie i międzymózgowie[19].
U ssaków z tyłomózgowia powstają 2 pęcherzyki, tworzące tyłomózgowie wtórne i rdzeniomózgowie (rdzeń przedłużony)[19].
Ewolucja mózgu ludzkiego
U przodków współczesnych ludzi w trakcie ewolucji dochodziło do stopniowego wzrostu objętości mózgu. Proces ten rozpoczął się od objętości około 600 cm3 u Homo habilis, osiągając 1500 cm3 u Homo sapiens neanderthalensis. Istnieje ogólna korelacja pomiędzy objętością mózgu a inteligencją. Jednakże współczesny Homo sapiens ma mniejszą objętość mózgu (1250 cm3) od neandertalczyka, a zwane hobbitami Homo floresiensis cechowały się objętością czaszki 380 cm3, około jednej trzeciej objętości Homo erectus, zbyt małej nawet jak dla szympansa. Zaproponowano, że H. floresiensis wyewoluował z H. erectus w mechanizmie karłowacenia wyspowego. Pomimo niewielkiego mózgu istnieją dowody na używanie przez hobbita ognia i wykorzystywanie kamiennych narzędzi, zbyt wysublimowanych dla proponowanego przodka, H. erectus[20]. Iain Davidson podsumował ewolucyjne ograniczenie wielkości mózgu, uznając, że powinien on być tak wielki, jak potrzeba, i tak mały, jak może być[21].
Przypisy
- 1 2 3 Hiller i Zegarska 2004 ↓, s. 42-43.
- ↑ Evolution of nervous systems. W: Gary G. Matthews: Neurobiology: molecules, cells, and systems. Wiley-Blackwell, 2001, s. 21. ISBN 0-632-04496-9. (ang.).
- ↑ Hiller i Zegarska 2004 ↓, s. 43.
- ↑ Sakarya O, Armstrong KA, Adamska M, Adamski M, Wang IF, Tidor B, Degnan BM, Oakley TH, Kosik KS. A post-synaptic scaffold at the origin of the animal kingdom. „PLoS ONE”. 2 (6), s. e506, 2007. DOI: 10.1371/journal.pone.0000506. PMID: 17551586. PMCID: PMC1876816.
- ↑ Jacobs DK1, Nakanishi N, Yuan D, et al.. Evolution of sensory structures in basal metazoa. „Integr Comp Biol”. 47 (5), s. 712–723, 2007. DOI: 10.1093/icb/icm094. PMID: 21669752.
- 1 2 3 Ruppert EE, Fox RS, Barnes RD: Invertebrate Zoology. Wyd. 7. Brooks / Cole, 2004, s. 111–124. ISBN 0-03-025982-7.
- ↑ Sanes DH, Reh TA, Harris WA: Development of the nervous system. Academic Press, 2006, s. 3–4. ISBN 978-0-12-618621-5. (ang.).
- ↑ Balavoine G. The segmented Urbilateria: A testable scenario. „Int Comp Biology”. 43 (1), s. 137–47, 2003. DOI: 10.1093/icb/43.1.137.
- ↑ Ghysen A. The origin and evolution of the nervous system. „Int. J. Dev. Biol.”. 47 (7–8), s. 555–62, 2003. PMID: 14756331. (ang.).
- ↑ Erwin DH, Davidson EH. The last common bilaterian ancestor. „Development”. 129 (13), s. 3021–32, lipiec 2002. PMID: 12070079.
- ↑ Bourlat SJ, Juliusdottir T, Lowe CJ, et al.. Deuterostome phylogeny reveals monophyletic chordates and the new phylum Xenoturbellida. „Nature”. 444 (7115), s. 85–8, November 2006. DOI: 10.1038/nature05241. PMID: 17051155.
- ↑ Lichtneckert R, Reichert H. Insights into the urbilaterian brain: conserved genetic patterning mechanisms in insect and vertebrate brain development. „Heredity”. 94 (5), s. 465–77, Maj 2005. DOI: 10.1038/sj.hdy.6800664. PMID: 15770230.
- 1 2 3 4 5 6 7 8 Hiller i Zegarska 2004 ↓, s. 45.
- 1 2 3 Adey WR. The nervous system of the earthworm Megascolex. „J. Comp. Neurol.”. 94 (1), s. 57–103, February 1951. DOI: 10.1002/cne.900940104. PMID: 14814220. (ang.).
- ↑ Wormbook: Specification of the nervous system.
- ↑ Ch. 20: Nervous system. W: Chapman RF: The insects: structure and function. Cambridge University Press, 1998, s. 533–568. ISBN 978-0-521-57890-5. (ang.).
- ↑ Chapman, s. 546
- ↑ Hiller i Zegarska 2004 ↓, s. 45-47.
- 1 2 3 Hiller i Zegarska 2004 ↓, s. 47.
- ↑ Brown P, Sutikna T, Morwood MJ, et al.. A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia. „Nature”. 431 (7012), s. 1055–61, 2004. DOI: 10.1038/nature02999. PMID: 15514638.
- ↑ Iain Davidson: As large as you need and as small as you can'--implications of the brain size of Homo floresiensis, (Iain Davidson). Une-au.academia.edu. [dostęp 2011-10-30].
Bibliografia
- Stanisław Hiller, Zofia Zegarska: Budowa ogólna śródmózgowia. W: Adam Bochenek, Michał Reicher, Wiesław Łasiński, Józef Markowski, Olgierd Narkiewicz: Anatomia człowieka. T. IV: Układ Nerwowy Ośrodkowy. Wydawnictwo Lekarskie PZWL, 2004, s. 219-220. ISBN 83-200-3449-3.
- kategoria
| Procesy ewolucyjne | |
|---|---|
| Nauki związane z ewolucją | |
| Rodzaje specjacji | |
| Rodzaje doboru naturalnego | |
| Ewolucja molekularna | |
| Ewolucja poszczególnych taksonów | |
| Pojęcia | |
| Historia myśli ewolucyjnej |
|
- Wikiprojekt:Biologia