| Arixeniidae | |||
| Jordan, 1909 | |||
 Arixenia esau | |||
| Systematyka | |||
| Domena | |||
|---|---|---|---|
| Królestwo | |||
| Typ | |||
| Gromada | |||
| Rząd | |||
| Podrząd |
Neodermaptera | ||
| Infrarząd |
Epidermaptera | ||
| Parvordo |
Eteodermaptera | ||
| Nadrodzina |
Forficuloidea | ||
| Rodzina |
Arixeniidae | ||
| Synonimy | |||
| |||
Arixeniidae – rodzina skorków z podrzędu Neodermaptera i nadrodziny Forficuloidea. Obejmuje 2 rodzaje i 5 opisanych gatunków. Żyją na guanie i skórze nietoperzy z podrodziny molosów (głównie naguska obrożnego). Są owłosione, całkowicie bezskrzydłe, o silnie zredukowanych oczach i nieprzekształconych w szczypce przysadkach odwłokowych. Występują w południowo-wschodniej części krainy orientalnej.
Taksonomia
Rodzina ta wprowadzona została w 1909 roku przez Karla Jordana jako monotypowa – z jednym rodzajem, Arixenia, opisanym przez tego samego autora w tej samej publikacji. Jordan zawarł sugestię, że rodzina ta może wymagać umieszczenia w osobnym podrzędzie[1], czego dokonał w 1911 Malcolm Burr, tworząc dla niej monotypowy podrząd Arixenina[2], którego pisownię potem zmieniono na Arixeniina zgodnie z zasadami języka łacińskiego[3]. Część autorów stosowała na przestrzeni lat inne klasyfikacje: Lucien Chopard w 1949 umieścił Arixeniidae w monotypowej nadrodzinie Arixenioidea, Anton Handlirsch w 1925 w podrzędzie Forficulina[3], a Edward Popham w 1965 w nadrodzinie Labioidea[4]. W 1974 T.C. Maa wyróżnił w obrębie rodziny nowy rodzaj Xeniaria[3]. Późniejsze analizy filogenetyczne wykazały, że podrząd Forficulina nie jest monofiletyczny, jeśli wyłączyć zeń Arixeniidae[5]. Nowsze, bliższe filogenezie podziały skorków uwzględniają Neodermaptera jako jedyny ich podrząd współczesny[6]. Rodzina Arixeniidae jest natomiast umieszczana w nadrodzinie Forficuloidea. Dotychczas opisano 5 gatunków z tej rodziny, zaliczanych do dwóch wymienionych wcześniej rodzajów[7].
Opis
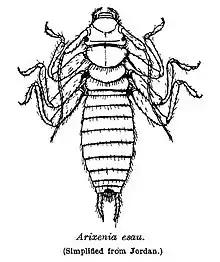
Skorki te mają umiarkowanie spłaszczone ciało, osiągające od 14 do 27 mm długości. Z wyjątkiem nieco jaśniejszych, małych, łysych znaków na głowie, tułowiu i udach oskórek jest prawie jednolicie porośnięty włoskami. Głowę cechuje silnie łukowaty i pośrodku zanikający szew epistomalny oraz formujące prawie ciągłą, prostą linię ramiona szwu czołowego. Silnie zredukowane i znacznie mniejsze od panewek czułkowych oczy buduje 50–90 fasetek. Czułki osobników dorosłych buduje 14 członów, przy czym drugi człon ich biczyka ma dwie łatki z porów zmysłowych. Długość czułków wynosi około ⅔ długości ciała. Aparat gębowy ma po dwa zęby szczytowe na żuwkach wewnętrznych oraz 2–3 zęby wierzchołkowe i brak lub jeden ząb środkowy na każdej żuwaczce[3].
Przedtułów ma bardzo duże przetchlinki. Pokrywy jak i skrzydła tylnej pary są całkowicie zanikłe. Długie i smukłe odnóża mają wszystkie uda podobnej długości i szerokości, golenie około 7 razy dłuższe niż szerokie, a przednie i tylne stopy z członami trzecimi krótszymi niż dwa poprzednie razem wzięte[3].
Odwłok samicy ma tergity ósmy i dziewiąty niezredukowane, sternit siódmy szeroko z tyłu zaokrąglony, sternit ósmy na większej powierzchni prawie błoniasty, sternit dziewiąty silnie zesklerotyzowany, a pygidium o szeroko zaokrąglonym lub trójkątnym i spiczastym wierzchołku. U samca manubrium oddzielone jest od symetrycznego z tyłu dziewiątego sternitu M-kształtnym lub podkowiastym paskiem. Włochate przysadki odwłokowe są u samicy smukłe, prawie walcowate i słabo zakrzywione, zaś u samca do silnego, kątowego zakrzywienia w nasadowej ⅓ lub około ½ długości tęgie, a dalej smukłe i prawie walcowate. Na każdej przysadce samca występują jedno lub dwa pólka papilli zmysłowych. Narządy genitalne samca cechują niewielkie, symetryczne, słabo zesklerotyzowane paramery, jeden lub cztery silnie zesklerotyzowane skleryty endofalliczne oraz położony w silnie zesklerotyzowanej części rurkowatej przewód wytryskowy[3].
Biologia i ekologia
_(15502213931).jpg.webp)
Przedstawiciele rodziny są związani z nietoperzami, głównie z bezwłosym naguskiem obrożnym (Cheiromeles torquatus), a okazjonalnie z moloskiem pomarszczonym (Tadarida plicata). Spotykani są wyłącznie w jaskiniach i dziuplach, zasiedlonych przez te nietoperze oraz na ich ciele. Związane są jednak wyłącznie z dużymi koloniami tych nietoperzy, zwykle położonymi wysoko nad powierzchnią gruntu[3]. Ich relację określa się w literaturze jako pasożytnictwo zewnętrzne[3][8] lub komensalizm[5]. Większość czasu skorki te spędzają na guanie zalegającym na ścianach i stropach grzęd nietoperzy, prawdopodobnie żerując na odżywiających się guanem stawonogach. Na ciała gospodarzy wchodzą prawdopodobnie celem pożywiania się na ich wydzielinach, choć samego żerowania nigdy nie zaobserwowano. Szczególnie chętnie wchodzą na osobniki martwe, umierające i młode. Obserwacje dotyczą jednak tylko młodych stadiów rozwojowych skorków, poukrywanych w fałdach skóry gospodarzy i nie jest jasne, czy starsze nimfy i osobniki dorosłe również wykazują takie zachowanie. Przypuszcza się, że przedstawiciele rodzaju Arixenia, jako że mają mniej szczecinek na ciele, więcej czasu spędzają na nietoperzach i ich wydzieliny stanowią większą część ich diety. Natomiast silniej porośnięte szczecinkami i mające silniejsze, przystosowane do przeżuwania żuwaczki gatunki z rodzaju Xeniaria więcej czasu spędzają na guanie, polując na zasiedlające je stawonogi. Ta niewielka rozbieżność w niszach tłumaczyłaby współwystępowanie przedstawicieli obu rodzajów na tych samych stanowiskach[3].
Arixeniidae są żyworodne. Zarodki odżywiane są w ciele samicy za pośrednictwem nibyłożyska (łac. pseudoplacenta). W rozwoju pozazarodkowym występują cztery stadia larwalne (nimfy), różniące się od stadium dorosłego m.in. rozmiarami oraz liczbą członów czułków. Nimfy mają 8 członów w I stadium, 11 członów w II, 12 w III oraz 13 w IV. Wzrost liczby członów czułków odbywa się przez podział członu trzeciego, czyli meristonu[3].
Rozprzestrzenienie
Zasięg tych skorków jest nierozerwalnie związany z zasięgiem ich gospodarzy. Obejmuje on region Półwyspu Malajskiego oraz wyspy Archipelagu Malajskiego w południowo-wschodniej części krainy orientalnej[3][8].
Przypisy
- ↑ K. Jordan. Description of a new kind of apterous earwig, apparently parasitic on a bat. „Novitates Zoologicae”. 16, s. 313-326, 1909.
- ↑ M. Burr. On some South African Dermaptera (Earwigs) in the South African Museum, Cape Town. „Annals of the South African Museum”. 10 (1), s. 1-17, 1911.
- 1 2 3 4 5 6 7 8 9 10 11 Satsuko Nakata, T.C. Maa. A review of the parasitic earwigs. „Pacific Insects”. 16, s. 307–374, 1974.
- ↑ E.J. Popham. A key to Dermaptera subfamilies (new classification of Dermaptera proposed). „Entomologist”. 98, s. 126-136, 1965.
- 1 2 K.J. Jarvis, F. Haas, M.F. Whiting. A phylogeny of earwigs (Insecta: Dermaptera) based on molecular and morphological evidence: reconsidering the classification of Dermaptera. „Systematic Entomology”. 30, s. 1–12, 2004.
- ↑ Dermaptera: The Earwins. W: David Grimaldi, Michael S. Engel: Evolution of the Insects. Cambridge University Press, 2005, s. 217–222.
- ↑ Heidi Hopkins, Michael D. Maehr, Fabian Haas, Lesley S. Deem: family Arixeniidae Jordan, 1909. [w:] Dermaptera Species File (Version 5.0/5.0) [on-line]. [dostęp 2019-01-06].
- 1 2 Piotr Naskręcki: rząd: skorki (cęgosze) – Dermaptera. W: Zoologia: Stawonogi. T. 2, cz. 2 Tchawkodyszne. Czesław Błaszak (red. nauk.). Warszawa: Wydawnictwo Naukowe PWN, 2012, s. 152-155.