| Allosaurus | |||
| Marsh, 1877 | |||
 | |||
| Systematyka | |||
| Domena | |||
|---|---|---|---|
| Królestwo | |||
| Typ | |||
| Podtyp | |||
| Gromada | |||
| Podgromada | |||
| Infragromada | |||
| Nadrząd | |||
| Rząd | |||
| Podrząd | |||
| Infrarząd | |||
| (bez rangi) | karnozaury | ||
| Nadrodzina | |||
| Rodzina | |||
| Rodzaj |
allozaur | ||
| Synonimy | |||
| |||
| Gatunki | |||
|
| |||
Allozaur (Allosaurus) – rodzaj dużego teropoda z rodziny allozaurów (Allosauridae) żyjącego w późnej jurze, 155–145 mln lat temu. Nazwa Allosaurus, oznaczająca „inny jaszczur”, pochodzi od greckich słów αλλος/allos („inny”, „dziwny”) oraz σαυρος/sauros („jaszczur”). Pierwsze szczątki z pewnością należące do przedstawicieli tego rodzaju zostały opisane w 1877 roku przez Othniela Charlesa Marsha. Jako jeden z pierwszych dobrze poznanych teropodów przez długi czas wzbudzał zainteresowanie kręgów pozapaleontologicznych i był dinozaurem często pojawiającym się w filmach fabularnych i dokumentalnych.
Allosaurus był dużym dwunożnym drapieżnikiem z wielką, wyposażoną w ostre zęby czaszką. Mierzył przeciętnie 8,5 m długości, jednak fragmentaryczne szczątki sugerują, że mógł osiągać ponad 12 m. W porównaniu z potężnymi tylnymi kończynami trójpalczaste kończyny przednie były niewielkie, a całe ciało równoważył długi, ciężki ogon. Allosaurus jest klasyfikowany w rodzinie Allosauridae, należącej do karnozaurów. Rodzaj ma skomplikowaną historię taksonomiczną i obejmuje nieokreśloną liczbę akceptowanych gatunków, spośród których najlepiej poznanym jest Allosaurus fragilis. Większość szczątków allozaurów pochodzi z północnoamerykańskiej formacji Morrison, materiał kopalny jest także znany z Portugalii i być może Tanzanii. Przez ponad połowę XX wieku był znany pod nazwą Antrodemus, jednak badania skamieniałości z Cleveland-Lloyd Dinosaur Quarry przywróciły nazwę Allosaurus i uczyniły allozaura jednym z najlepiej poznanych dinozaurów.
Jako główny duży drapieżnik formacji Morrison Allosaurus stał na szczycie łańcucha pokarmowego, prawdopodobnie polując na duże dinozaury roślinożerne, takie jak ornitopody, stegozaury i zauropody. Niektórzy paleontolodzy uważają, że allozaur prowadził stadny tryb życia i polował w grupach, podczas gdy inni twierdzą, iż dorosłe osobniki były wobec siebie agresywne, a zgromadzenia dużej ilości szczątków przedstawicieli tego rodzaju w jednym miejscu dowodzą, że samotne zwierzęta pożywiały się tą samą zdobyczą.
Budowa

Allosaurus był zbudowany w sposób typowy dla dużych teropodów – miał masywną czaszkę osadzoną na krótkiej szyi, długi ogon i zredukowane kończyny przednie. Allosaurus fragilis, najlepiej poznany i jedyny pewny gatunek, mierzył przeciętnie 8,5 m długości[3], podczas gdy długość największego okazu należącego bezdyskusyjnie do tego gatunku (AMNH 680) szacuje się na 9,7 m[4], a jego masę na 2,3 t[4]. Jednak w świetle najnowszych badań, do Allosaurus fragilis należy także wyróżniany niegdyś, znany z ogromnego, niekompletnego szkieletu rodzaj Epanterias, co świadczy, że w pełni dojrzałe allozaury osiągały ponad 12 m długości[1][2]. Już James Madsen, w monografii z 1976, wspomina o kościach wskazujących na maksymalną długość dochodzącą do 12–13 metrów[5]. Jak w przypadku większości dinozaurów oszacowania masy są niepewne; od 1980 roku wynoszą od 1000 do 4000 kg, z 1010 kg jako dominantą[1]. John Foster, specjalista od formacji Morrison, sugeruje, że masa 1000 kg jest prawdopodobna dla dużego dorosłego Allosaurus fragilis, jednak 700 kg jest bardziej odpowiednie dla osobnika średniej wielkości, którego kości udowe mierzył[6]. Analizy wykorzystujące trójwymiarowy model „Wielkiego Ala” (patrz niżej) wskazują, że osobnik ten – niebędący jeszcze w pełni dorosły – ważył najprawdopodobniej około 1500 kg, co sugeruje, że dojrzałe zwierzęta osiągały jeszcze większą masę[7].
Kilka olbrzymich okazów przypisywanych allozaurowi może w rzeczywistości być przedstawicielami innych rodzajów. Blisko spokrewniony rodzaj Saurophaganax (OMNH 1708) dorastał prawdopodobnie do 10,9 m długości[4], a jego jedyny gatunek bywał niekiedy klasyfikowany jako Allosaurus maximus, jednak nowsze badania potwierdzają odrębność Saurophaganax od Allosaurus[8]. Innym potencjalnym allozaurem jest duży teropod, znany z niekompletnego szkieletu odkrytego w Peterson Quarry w osadach formacji Morrison w Nowym Meksyku; ten duży allozauryd może być kolejnym osobnikiem zaurofaganaksa[9]. Do Allosaurus należy natomiast prawdopodobnie opisany przez Cope'a Epanterias (AMNH 5767), mierzący prawdopodobnie ok. 12,1 m długości[4], co świadczyło by o tym, że allozaur był w rzeczywistości znacznie większy niż sądzono. Hipotezę tą popierają najnowsze badania[1][2].
Czaszka

Czaszka i zęby allozaura były umiarkowanej wielkości jak na teropoda jego rozmiarów. Paleontolog Gregory Paul ocenia, że długość czaszki osobnika mierzącego około 7,9 m długości wynosiła 84,5 cm[10]. W obu kościach przedszczękowych znajdowało się po pięć D-kształtnych w przekroju zębów, zaś w każdej kości szczękowej – od czternastu do siedemnastu; ich liczba nie odpowiadała dokładnie rozmiarowi kości. Kości zębowe także zawierały 14–17 zębów, średnio 16. Zęby stawały się krótsze, węższe i bardziej zakrzywione, im bliżej tyłu szczęk się znajdowały. Wszystkie miały piłkowane krawędzie. Zęby łatwo wypadały, będąc zastępowane kolejnymi, co czyni z nich pospolite skamieniałości[5].
Nad i przed oczami znajdowała się para rogów, będących przedłużeniem kości łzowych i zróżnicowanych pod względem kształtu i rozmiaru. Różki były prawdopodobnie pokryte keratynową pochwą i mogły pełnić różne funkcje, takie jak ochrona przed Słońcem[5], ozdoba lub broń służąca do walk z innymi przedstawicielami tego samego gatunku[10][11] (mimo ich kruchości[5]). Garb z tyłu czaszki służył jako miejsce przyczepu mięśni, podobnie jak u tyranozaurów[11].
Wewnątrz kości łzowych znajdowały się wgłębienia mogące mieścić gruczoły, takie jak gruczoły solne[12]. Kości szczękowe miały zatoki szczękowe wykształcone lepiej niż u bardziej bazalnych teropodów, takich jak ceratozaur czy marszozaur. Może to mieć związek ze zmysłem węchu; możliwe, że allozaur był wyposażony w narząd przypominający narząd Jacobsona. Szczyt czaszki był cienki, prawdopodobnie w celu poprawienia termoregulacji mózgu[5]. Czaszka i żuchwa miały stawy pozwalające na ruch kości. Kości przedniej i tylnej połowy żuchwy łączyły się z sobą luźno, umożliwiając szczękom wyginanie się na zewnątrz i zwiększanie kąta ich rozwarcia[13]. Nie można także wykluczyć obecności stawów w obrębie mózgoczaszki i kości czołowych[5].

Szkielet pozaczaszkowy
Szyja allozaura składała się z dziewięciu kręgów, grzbiet z czternastu, a kość krzyżowa, wspierająca biodra – z pięciu[5]. Liczba kręgów ogonowych jest nieznana i zależna od rozmiarów osobnika; James Madsen ocenia ich liczbę na około 50[5], podczas gdy Gregory S. Paul uznaje, że było ich nie więcej niż 45[10]. W szyi oraz na przedzie kręgów grzbietowych znajdowały się wydrążone przestrzenie[5]. Uważa się, że przestrzenie te, znajdowane także u innych teropodów (w tym ptaków), mieściły worki powietrzne służące do oddychania[2]. Klatka piersiowa była szeroka, bardziej beczkowata niż u mniej zaawansowanych teropodów, takich jak ceratozaur[14]. Gastralia (żebra brzuszne) allozaura rzadko zachowują się w zapisie kopalnym, przeważnie w złym stanie[10]. W jednym znanym przypadku gastralia są dowodem na uraz nabyty za życia zwierzęcia[15]. Widełki obojczykowe również występowały u allozaura, choć rozpoznano je dopiero w 1996 roku – wcześniej mylone były z żebrami brzusznymi[8][16]. Kość biodrowa, będąca główną kością stawu biodrowego, była masywna, a kość łonowa zakończona stopką, mogącą służyć jako miejsce przyczepu silnych mięśni lub do podpierania ciała podczas odpoczynku na ziemi. Madsen spostrzegł, że u około połowy spośród osobników znalezionych w Cleveland-Lloyd Dinosaur Quarry, niezależnie od rozmiarów, kości łonowe nie były ze sobą połączone od strony stopki. Zasugerował, że jest to cecha odróżniająca samice od samców. Jego zdaniem kości łonowe samic nie łączyły się ze sobą, co miało ułatwiać składanie jaj[5]. Hipoteza ta nie spotkała się jednak z powszechną akceptacją.

Kończyny przednie allozaura były krótkie w porównaniu do tylnych (35% długości kończyn tylnych u osobników dorosłych). Miały po trzy palce, zakończone dużymi, silnie zakrzywionymi i ostrymi pazurami[5]. Ramiona były potężne, a przedramiona nieco od nich krótsze[10] (stosunek długości kości ramiennej do łokciowej wynosił 1:1,2)[17]. W nadgarstku znajdowała się odmiana kości księżycowatej[18] znajdowanej także u bardziej zaawansowanych teropodów, takich jak maniraptory. Z trzech palców wewnętrzny (kciuk) był największy[10] i odmienny od pozostałych dwóch[17]. Nogi nie były tak długie i przystosowane do dużych prędkości jak u przedstawicieli Tyrannosauridae, a pazury nie były tak wykształcone i bardziej przypominały kopyta niż u wcześniejszych teropodów[10]. Każda stopa miała trzy palce podporowe oraz tylny pazur, nie pełniący takich funkcji. Według Madsena mógł on być wykorzystywany do chwytania młodych[5]. Niektórzy sugerują istnienie piątego palca (najbardziej zewnętrznego) pełniącego funkcję dźwigni pomiędzy ścięgnem Achillesa a stopą[19].
Klasyfikacja

Allozaur należał do Allosauridae, rodziny dużych teropodów z grupy karnozaurów. Nazwa ta została ustanowiona dla tego rodzaju w 1878 przez Othniela Charlesa Marsha[20], jednak była rzadko używana aż do lat 70. XX wieku, ustępując Megalosauridae, innej rodziny dużych teropodów, stanowiącej wówczas „takson-kosz na śmieci” (takson, do którego włączano większość gatunków dinozaurów drapieżnych o niepewnej klasyfikacji). W publikacjach z tamtego okresu, aż do monografii Jamesa Madsena z 1976, nie pojawia się również nazwa Allosaurus, lecz Antrodemus. Głównymi autorami używającymi nazwy Megalosauridae zamiast Allosauridae byli: Gilmore, 1920[17]; von Huene, 1926[21]; Romer, 1956[22] i 1966[23]; Steel, 1970[24] oraz Walker, 1964[25].
Praca Madsena sprawiła, że nazwa Allosauridae stała się preferowana na określenie tej rodziny. Grupa ta jednak nie została precyzyjnie zdefiniowana. Prace popularnonaukowe zaliczały do niej różne rodzaje dużych teropodów, przeważnie większych i lepiej poznanych niż megalozaury. Typowymi teropodami uważanymi za blisko spokrewnione z allozaurem były: indozaur, piatnickizaur, piwetozaur, jangczuanozaur[26], akrokantozaur, czilantajzaur, kompsozuch, stoksozaur i syczuanozaur[27]. Nowsze badania oraz ustalanie pokrewieństwa ewolucyjnego za pomocą analiz kladystycznych spowodowało, że żaden z wymienionych dinozaurów oprócz akrokantozaura (którego bliższe pokrewieństwo jest niejasne), jangczuanozaura[2] i czilantajzaura[28] nie jest zaliczany do allozauroidów (Allosauroidea).
Allozaury to jedna z czterech rodzin karnozaurów; pozostałe trzy to karcharodontozaury (Carcharodontosauridae), sinraptory (Sinraptoridae) i Neovenatoridae[28]. Przedstawiciele Allosauridae w pracy Gregory'ego Paula Predatory Dinosaurs of the World zostali uznani za przodków tyranozaurów (grupa allozaury-tyranozaury byłaby polifiletyczna)[29], jednak koncepcja ta została odrzucona, gdyż tyranozaury są uznawane za członków odrębnej linii ewolucyjnej teropodów – celurozaurów[30]. Allosauridae to najmniejsza rodzina karnozaurów – obejmuje, oprócz allozaura, jedynie zaurofaganaksa i nienazwanego jeszcze allozauroida z Francji[2]. Inny rodzaj, Epanterias, jest kolejnym hipotetycznie ważnym członkiem grupy, jednak zarówno on jak i zaurofaganaks mogą okazać się jedynie dużymi osobnikami Allosaurus[10]. Najnowsze badania utrzymują jednak rodzaj Saurophaganax, a włączają Epanterias do Allosaurus[1][2]. Według większości obecnych analiz akrokantozaur należy do karcharodontozaurów[31][2][32], aczkolwiek niektórzy naukowcy umieszczają go w Allosauridae[33].
Historia odkryć
Wczesne odkrycia i badania
Odkrycie i pierwsze badania allozaura utrudniała ilość nazw ukutych w trakcie wojny o kości pod koniec XIX wieku. Pierwszą opisaną w trakcie konfliktu skamieniałością była kość nabyta z drugiej ręki przez Ferdinanda Vandiveera Haydena w 1869 roku. Pochodziła ona z Middle Park w pobliżu Granby w stanie Kolorado, prawdopodobnie ze skał formacji Morrison. Mieszkańcy tamtych terenów identyfikowali podobne kości jako „skamieniałe końskie kopyta”. Hayden przesłał swoje znalezisko Josephowi Leidy'emu, który zidentyfikował je jako połowę kręgu ogonowego i wstępnie przypisał do europejskiego rodzaju Poekilopleuron jako Poicilopleuron valens [sic!][34]. Później uznał, że kość zasługuje na osobny rodzaj – Antrodemus[35].
Allosaurus został opisany w oparciu o YPM 1930, niewielki zbiór niekompletnych kości zawierający trzy kręgi, fragment żebra, kość stopy oraz prawą kość ramienną, która okazała się najprzydatniejsza w późniejszych dyskusjach. Othniel Charles Marsh nadał tym szczątkom formalną nazwę Allosaurus fragilis w 1877 roku. Allosaurus pochodzi od greckich słów: allos/αλλος – oznaczającego „inny” lub „dziwny” – oraz sauros/σαυρος („jaszczur”, „gad”)[36]. Otrzymał swą nazwę ze względu na budowę kręgów odmienną od innych znanych wówczas dinozaurów[37][38]. Nazwa gatunkowa fragilis (łac. „kruchy”) odnosi się do lekkości kręgów. Kości zostały wydobyte ze skał formacji Morrison w pobliżu Garden Park, na północ od Cañon City[37]. Marsh i Edward Drinker Cope, rywalizujący ze sobą, w oparciu o zbliżony materiał kopalny ustanowili kilka innych rodzajów, które później figurowały w historii taksonomicznej Allosaurus. Są to opisane przez Marsha Creosaurus[20] i Labrosaurus[39] oraz Epanterias Cope'a[40].



Pracując w pośpiechu Marsh i Cope nie zawsze śledzili odkrycia dokonywane przez ich zespoły. Po odkryciu przez Benjamina Mudge'a holotypu Allosaurus Marsh zdecydował się skoncentrować na wykopaliskach w Wyoming. Po wznowieniu prac w Garden Park w 1883 roku M. P. Felch odnalazł jeden niemal kompletny i kilka fragmentarycznych szkieletów[12]. Jeden z członków ekipy Cope'a, H. F. Hubbell, w 1879 roku w Como Bluff odkrył szkielet allozaura, jednak prawdopodobnie nie wspomniał o jego kompletności, gdyż Cope nigdy nie zbadał skamieniałości. Po przeanalizowaniu materiału w 1903 (sześć lat po śmierci Cope'a) został uznany za jeden z najbardziej kompletnych znanych szkieletów teropodów. W 1908 roku, skatalogowany jako AMNH 5753, został wystawiony w Amerykańskim Muzeum Historii Naturalnej[41]. Zmontowany w pozycji imitującej pożywianie się apatozaurem, mimo iż jest pierwszym wystawionym na widok opinii publicznej, a także często ilustrowanym oraz fotografowanym szkieletem, nie został nigdy naukowo opisany[42].
Późniejsze badania utrudniała wielość nazw utworzonych w lakonicznych opisach Marsha i Cope'a. Już autorzy tacy jak Samuel Wendell Williston sugerowali, że ukutych zostało zbyt wiele nazw[43]. Williston w 1901 roku stwierdził, że Marsh nigdy nie był w stanie rozróżnić Allosaurus i Creosaurus[44]. Najbardziej znaczącą próbę uproszczenia skomplikowanej sytuacji podjął w 1920 roku Charles Whitney Gilmore. Doszedł on do wniosku, że kręg ogonowy Antrodemus był nieodróżnialny od kręgu allozaura, a preferowaną nazwą powinien stać się Antrodemus, gdyż jako starsza nazwa miał priorytet[17]. Antrodemus stał się dominującym określeniem na ponad pół wieku, do momentu publikacji Jamesa Madsena opisującej osobniki z Cleveland-Lloyd, w której stwierdził on, że Allosaurus jest bardziej właściwą nazwą, gdyż Antrodemus opierał się o materiał niedający informacji o cechach diagnostycznych rodzaju oraz miejscu pochodzenia (nie jest znana np. formacja geologiczna, w której znaleziono pojedynczą kość Antrodemus)[5]. Nazwa „Antrodemus” bywa używana dla wygody w celu odróżnienia czaszki opisanej przez Gilmore'a od czaszki zrekonstruowanej przez Madsena[45].
Odkrycia w Cleveland-Lloyd
Mimo iż sporadyczne prace w miejscu znanym później jako Cleveland-Lloyd Dinosaur Quarry w Emery County w stanie Utah miały miejsce już w 1927 roku, a stanowisko paleontologiczne zostało opisane w 1945 przez Williama J. Stokesa[46], główne operacje nie rozpoczęły się aż do 1960. W wyniku wspólnych prac ponad 40 instytucji pomiędzy 1960 a 1965 wydobyto tysiące kości[5]. Stanowisko to jest znane z dominacji szczątków Allosaurus fragilis oraz braku jasnego stanowiska naukowców, co jest tego przyczyną. Spośród kości co najmniej 73 różnych dinozaurów przynajmniej 46 należy do allozaurów. Odnalezione tam kości były zdezartykulowane i wymieszane. Opublikowano około tuzina prac naukowych o tafonomii cmentarzyska, w których prezentowano sprzeczne stanowiska na temat przyczyn jego powstania. Sugerowano śmierć zwierząt w wyniku ugrzęźnięcia na torfowisku, w głębokim błocie, z powodu suszy, gdy zwierzęta zgromadziły się wokół wysychającego akwenu, a także utonięcia we wzbierającym stawie lub bagnie[47]. Niezależnie od faktycznych przyczyn powstania cmentarzyska, ogromna liczba dobrze zachowanych kości uczyniła z allozaura jednego z najlepiej poznanych rodzajów teropodów. W kamieniołomie odnalezione zostały szkielety dinozaurów należących do niemal wszystkich grup wiekowych i zróżnicowanych pod względem rozmiarów – od osobników mierzących mniej niż metr długości[48] do teropodów ponad dwunastometrowych. Dezartykulacja szczątków jest pomocna przy ich opisywaniu, gdyż zwykle odnajdowane są połączone[5].
Badania po 1980
W okresie od wydania monografii Madsena badania nad allozaurem dotyczyły głównie jego paleobiologii i paleoekologii. Publikowano prace dotyczące zmienności w budowie szkieletu allozaurów[49], wzrostu[50][51], budowy czaszki[52] i mózgu[53], metod polowania[54] oraz możliwości prowadzenia stadnego trybu życia i opieki rodzicielskiej[55]. Ponowna analiza wcześniej odkrytych skamieniałości (zwłaszcza dużych osobników)[10][56], odkrycia w Portugalii[57] oraz kilka nowych, bardzo kompletnych okazów[8][58][59], także przyczyniły się do zwiększenia wiedzy o allozaurach.
Wielki Al

Jednym z najbardziej znaczących znalezisk allozaura było odkrycie w 1991 „Wielkiego Ala” (MOR 693), niemal w 95% kompletnego szkieletu osobnika mierzącego w chwili śmierci około 8 m długości. MOR 693 został wydobyty niedaleko Shell w Wyoming przez ekipę Muzeum Gór Skalistych (Museum of the Rockies) oraz Muzeum Geologicznego Uniwersytetu Wyoming (University of Wyoming)[60]. Szkielet ten został odkryty przez szwajcarski zespół prowadzony przez Kirby'ego Sibera. Ta sama ekipa wydobyła później drugiego allozaura, „Wielkiego Ala Dwa”, który do dziś jest najlepiej zachowanym szkieletem tego typu[59].
Do nadania „Wielkiemu Alowi” jego imienia przyczyniły się stopień zachowania, kompletność oraz znaczenie naukowe jego szkieletu – sam osobnik nie osiągnął przeciętnej wielkości Allosaurus fragilis[60] (choć niektórzy sugerowali, że może należeć do odrębnego gatunku[7]; Chure i Loewen (2020) zaliczyli go do gatunku A. jimmadseni[61]) i był podrostkiem, którego poziom rozwoju w stosunku do dorosłego szacuje się na 87%[62]. Został on opisany w 1996 roku przez Brenta Breithaupta[63]. Dziewiętnaście z jego kości było złamanych lub nosiło ślady infekcji, które mogły przyczynić się do jego śmierci. Dotknięte patologiami było pięć żeber, pięć kręgów i cztery kości stopy; kilka uszkodzonych kości nosi ślady osteomyelitis, zapalenia kości. Głównym problemem za życia była dla „Wielkiego Ala” infekcja prawej stopy, wpływająca prawdopodobnie na zdolność do poruszania się i mogąca prowadzić do kontuzji drugiej stopy ze względu na zmianę sposobu chodzenia[62].
Gatunki i taksonomia

Nie jest jasne, jak wiele gatunków należało do rodzaju Allosaurus. Od 1988 za potencjalnie ważne uznawano siedem gatunków (A. amplexus[10], A. atrox[10], A. europaeus[64], gatunek typowy A. fragilis[2], A. jimmadseni[8][61], A. maximus[49] i A. tendagurensis[2]), jednak tylko niektóre z nich były akceptowane w danym momencie. Ponadto na przestrzeni lat co najmniej dziesięć wątpliwych lub nieopisanych gatunków włączano do Allosaurus, wśród nich także te należące do rodzajów uznawanych obecnie za jego synonimy. W pracy dotyczącej bazalnych tetanurów w drugim wydaniu The Dinosauria tylko Allosaurus fragilis (w tym A. amplexus i A. atrox jako synonimy), A. jimmadseni (wówczas jeszcze nieznawany) i A. tendagurensis były uwzględniane jako potencjalnie ważne. A. europaeus nie był wówczas opisany, a A. maximus przypisano do rodzaju Saurophaganax[2].
Wszystkie skamieniałości A. amplexus, A. atrox, A. fragilis, A. jimmadseni i A. maximus są znane z datowanych na kimeryd lub tyton osadów formacji Morrison. A. fragilis jest spośród nich najpospolitszy – znane są szczątki co najmniej sześćdziesięciu osobników[2]. Od końca lat 80. XX wieku trwa debata nad możliwością występowania tam dwóch gatunków allozaura – drugim, obok A. fragilis, miałby być A. atrox[10][65]. Nowsze badania wspierają hipotezę mówiącą o jednym gatunku, różnice w materiale kopalnym przypisując zmienności osobniczej[66][67]. Chure i Loewen (2020) przenieśli jednak szereg okazów odkrytych na obszarze stanów Kolorado, Utah i Wyoming do odrębnego gatunku A. jimmadseni, różniącego się od A. fragilis m.in. proporcjonalnie wyższymi rogami wyrastającymi z kości łzowych oraz występowaniem na kościach nosowych niskich kostnych grzebieni ciągnących się nieprzerwanie od kości przedszczękowych do kości łzowych[61].
A. europaeus, odnaleziony w kimerydzkich osadach portugalskiej formacji Lourinhã[64], może być identyczny z A. fragilis[68]. Cechy uznawane za wyjątkowe dla tego gatunku mogą występować również u A. fragilis lub być błędnie zinterpretowane, dlatego niektórzy autorzy uznają A. europaeus za nomen dubium, przynajmniej dopóki holotyp nie zostanie dokładniej opisany[69]. Szczątki A. tendagurensis, które odkryto w Tanzanii, datuje się na kimeryd[70]. Mimo iż w nowszych badaniach uznawany jest za ważny gatunek allozaura, mógł być bazalnym tetanurem[71] lub wątpliwym gatunkiem teropoda[3]. Mimo iż słabo poznany, był dużym teropodem, mierzącym około 10 m długości i ważącym 2,5 t[4].
Allosaurus jest uznawany za prawdopodobny synonim rodzajów Antrodemus, Creosaurus, Epanterias i Labrosaurus[2]. Większość gatunków synonimizowanych z A. fragilis lub błędnie przypisanych do tego rodzaju bazowanych jest na źle zachowanym materiale kopalnym. Wyjątkiem jest Labrosaurus ferox, nazwany w 1884 przez Marsha na podstawie nietypowo uformowanej, niekompletnej żuchwy, z widoczną luką w uzębieniu na czubku pyska, a bardzo szeroką w tylnej części[72]. Późniejsze badania dowiodły, że była to pozostałość po kontuzji odniesionej za życia zwierzęcia[17], a niespotykany kształt tylnej części czaszki spowodowany był spowodowany gipsową rekonstrukcją[73]. Obecnie żuchwa ta jest uważana za pozostałość osobnika z gatunku Allosaurus fragilis[2]. Doniesienia o skamieniałościach przypisywanych allozaurowi pochodziły z wielu rejonów, m.in. Australii[74], Syberii[75] oraz Szwajcarii[3], jednak szczątki te zostały uznane za należące do innych dinozaurów.
Paleoekologia
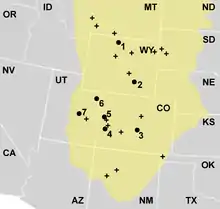
Allosaurus był najpospolitszym dużym teropodem na terenie zachodnioamerykańskiej formacji Morrison – 70–75% odnalezionych osobników to allozaury[6] – i jako taki był dominującym drapieżnikiem w tamtejszych ekosystemach[76]. W późnej jurze formacja Morrison prawdopodobnie była półpustynią z odrębnymi porami: suchą i deszczową oraz równinami zalewowymi[77]. Roślinność wahała się od nadrzecznych lasów iglastych, paproci i paproci drzewiastych do paprociowych sawann z rzadko rosnącymi drzewami[78].
Formacja Morrison obfituje w skamieniałości zielenic, grzybów, mchów, skrzypów, paproci, sagowców, miłorzębów oraz przedstawicieli kilku rodzin iglastych. Odkryto tam także szczątki małżów, ślimaków, ryb promieniopłetwych, płazów bezogonowych i ogoniastych, żółwi, sfenodontów, jaszczurek, lądowych i wodnych krokodylomorfów, kilku gatunków pterozaurów, bardzo licznych dinozaurów oraz wczesnych ssaków, takich jak dokodonty, wieloguzkowce i trykonodonty. Do dinozaurów znanych z osadów formacji Morrison należą teropody Ceratosaurus, Ornitholestes i Torvosaurus, zauropody Apatosaurus, Brachiosaurus, Camarasaurus i Diplodocus oraz ptasiomiedniczne Camptosaurus, Dryosaurus i Stegosaurus[79]. Późnojurajskie formacje w Portugalii, gdzie także występował Allosaurus, są uważane za podobne do formacji Morrison, jednak z silniejszym wpływem oceanicznym. Wiele dinozaurów z Ameryki Północnej występowało także w Portugalii lub miało tam swoich odpowiedników (Brachiosaurus i Lusotitan, Camptosaurus i Draconyx)[80].

Allosaurus, podobnie jak inne duże teropody, Ceratosaurus i Torvosaurus, występował zarówno na terenie obecnych Stanów Zjednoczonych, jak i Portugalii[80]. Badania anatomiczne oraz skamieniałości odkrytych w danej lokalizacji sugerują, że te trzy dinozaury zajmowały różne nisze ekologiczne. Ceratozaury i torwozaury mogły być bardziej aktywne w okolicach zbiorników wodnych, miały też niższe, szczuplejsze ciała, co dawało im przewagę w lasach i terenach gęsto zarośniętych, podczas gdy allozaury były masywniej zbudowane, z dłuższymi nogami, szybsze, lecz mniej zwrotne i prawdopodobnie preferowały suche równiny zalewowe[81]. Allosaurus sam także stanowił potencjalne pożywienie dla innych drapieżników, czego dowodzą ślady zębów na stopce kości łonowej allozaura, pozostawione prawdopodobnie przez ceratozaura lub torwozaura. Położenie kości w ciele (wzdłuż dolnej granicy piersi, częściowo osłonięta nogami) oraz fakt, że jest to jedna z najbardziej masywnych kości, dowodzi, że drapieżnik pożywiał się padliną allozaura[82].
Paleobiologia
Ontogeneza
Obfitość skamieniałości allozaurów z niemal wszystkich grup wiekowych pozwala naukowcom badać schemat ich wzrostu oraz tryb życia. Pokruszone skorupki jaj odnalezione w Kolorado są przypisywane przedstawicielom tego rodzaju[3]. Na podstawie histologicznej analizy kości kończyn górną granicę wieku Allosaurus szacuje się na 22 do 28 lat, co jest porównywalne z szacowaną długością życia innych dużych teropodów, takich jak tyranozaur. Te same badania wskazują, że maksymalne rozmiary były osiągane około 15. roku życia, a przyrost masy wynosił w przybliżeniu 150 kg rocznie[50].
Tkanka rdzenna, odnajdowana u zróżnicowanych dinozaurów, takich jak Tyrannosaurus i Tenontosaurus, została odnaleziona u co najmniej jednego osobnika w kości piszczelowej odkrytej w Cleveland-Lloyd Dinosaur Quarry. Obecnie tkanki tego typu występują jedynie u samic ptaków w okresie rozrodczym, które wykorzystują je w procesie wytwarzania skorupki jaja. Obecność tych tkanek wskazuje płeć zwierzęcia i sugeruje, że osiągnęło ono zdolność reprodukcyjną. Pierścienie przyrostowe kości wykazały, że osobnik ten zmarł w wieku dziesięciu lat, co dowodzi, że zwierzę osiągało dojrzałość płciową wcześniej niż maksymalne rozmiary[83].
Odkrycie szczątków młodocianego osobnika z niemal kompletnymi kończynami przednimi dowodzi, że kończyny dolne były stosunkowo dłuższe w młodości, a goleń i stopa – stosunkowo dłuższe niż udo. Różnice te sugerują, że młodsze allozaury były szybsze i miały odmienne strategie łowieckie niż dorosłe – prawdopodobnie ścigały niewielkie kręgowce, z wiekiem stając się polującymi z zasadzki dominującymi drapieżnikami[51]. W miarę wzrostu zwierzęcia kość udowa stawała się grubsza, szersza i mniej okrągła w przekroju, mięśnie stawały się krótsze, a wzrost kończyny zwalniał. Zmiany te dowodzą, że kończyny tylne osobników młodocianych były poddawane mniej przewidywalnym naciskom[84].
Odżywianie

Paleontolodzy uznają allozaura za aktywnego drapieżnika polującego na duże zwierzęta. Prawdopodobnie zarówno polował na zauropody, jak i żywił się martwymi osobnikami, czego dowodzą ślady zębów na kościach zauropodów oraz zgubione w ich pobliżu zęby allozaurów[85]. Są również dowody na ataki allozaurów na stegozaury, takie jak kręg ogonowy allozaura, częściowo wygojony po ranie zadanej kolcem ogonowym stegozaura, oraz U-kształtny ślad ukąszenia na płycie szyjnej stegozaura, odpowiadający kształtem pyskowi allozaura[86]. Jednakże, jak zauważył Gregory Paul w 1988, Allosaurus prawdopodobnie nie był zdolny do samotnego polowania na dorosłe zauropody ze względu na średniej wielkości głowę, stosunkowo niewielkie zęby i wielką dysproporcję rozmiarów pomiędzy allozaurem a zauropodem[10]. Możliwe również, że polował na osobniki młodociane, a nie w pełni dojrzałe[6][65]. Badania z lat 90. XX wieku i początków XXI wieku pozwoliły sformułować alternatywne hipotezy. Robert Bakker, porównując allozaura i kenozoiczne szablozębne ssaki, odnalazł kilka podobnych adaptacji, takich jak redukcja mięśni szczęk i rozwój mięśni szyi, a także zdolność do niezwykle szerokiego otwierania szczęk. Ponieważ allozaur nie miał przypominających szable zębów, Bakker zasugerował inny rodzaj ataku, wykorzystujący adaptacje szyi i szczęk. Stosunkowo niewielkie zęby pełniły w efekcie rolę ząbkowania – przypominającego ząbkowanie piły – ciągnącego się wzdłuż szczęki. Taki typ budowy szczęk pozwalał na wykonywanie licznych cięć powodujących wykrwawienie się ofiary[54].

Badania czaszki allozaura wykorzystujące metodę elementów skończonych doprowadziły do podobnych konkluzji. Analiza biomechaniczna wykazała, że czaszka była bardzo silna, lecz ze stosunkowo niewielką siłą ugryzienia. Przy wykorzystaniu wyłącznie mięśni siła ukąszenia wynosiła od 805 do 2148 niutonów – mniej niż w przypadku aligatorów (13 000 N), lwów (4167 N), lampartów (2268 N) – jednak czaszka była w stanie wytrzymać pionowy nacisk na zęby o sile 55 000 N. Rayfield i współpracownicy sugerują, że Allosaurus używał czaszki jak topora, atakując ofiary otwartą paszczą, tnąc mięso zębami i wyrywając jego kawały nie łamiąc kości – w przeciwieństwie do tyranozaura, o którym uważa się, że był zdolny do gruchotania kości. Sugerują także, że budowa czaszki umożliwiała allozaurowi stosowanie różnej techniki ataku w zależności od ofiary – była ona wystarczająco lekka, by atakować małe i zwinne ornitopody oraz wystarczająco mocna, by wytrzymać uderzenie dużej zdobyczy, takiej jak stegozaury i zauropody[52]. Interpretacja ta była podważana przez innych badaczy, którzy nie znaleźli współczesnej analogii do ataku szczękami niczym toporem i za bardziej prawdopodobne uznali, że czaszka była wytrzymała, aby zrekompensować jej otwartą konstrukcję podczas walki ze zdobyczą[87]. Rayfield i współpracownicy stwierdzili, że Allosaurus rzeczywiście nie ma współczesnych odpowiedników, jednak jego zęby były przystosowane do tego typu ataków, a artykulacje czaszki, uznane przez Frazzettę i Kardonga za problematyczne, faktycznie mogły pomagać w ochronie podniebienia i ograniczaniu nacisków[88]. Możliwe również, że teropody takie jak Allosaurus wyrywały kawały mięsa z ciał żywych zwierząt i nie musiały wysilać się na zabijanie zdobyczy. Dawało to także możliwość ponownego pożywienia się tą samą ofiarą po jej wyzdrowieniu[2]. Dodatkowo zasugerowano, że ornitopody były najpowszechniejszą możliwą zdobyczą, a allozaury polowały na nie w podobny sposób jak obecnie duże kotowate – chwytały ofiarę kończynami przednimi, a następnie wykonywały serię ugryzień w szyję, by zmiażdżyć tchawicę[6]. Jest to zgodne z innymi dowodami, wskazującymi że kończyny przednie allozaura były wystarczająco silne do przytrzymania zdobyczy[18].
Inne aspekty odżywiania obejmują budowę oczu, ramion i kończyn tylnych. Kształt czaszki allozaura ograniczał zdolność widzenia stereoskopowego do około 20° szerokości – nieco mniej niż w przypadku współczesnych krokodyli. Podobnie jak u nich zdaje się to wystarczać do ocenienia odległości od łupu i momentu ataku[89]. Podobne pole widzenia sugeruje, że allozaury, podobnie jak krokodyle, polowały z zasadzki[90]. Ramiona, w porównaniu z innymi teropodami, były przystosowane zarówno do trzymania ofiary na dystans, jak i przy ciele[18], a artykulacja pazurów sugeruje, że mogły być one używane jako haki[17]. Podobnie jak u innych nieptasich teropodów, środek ciężkości był zlokalizowany przed i poniżej stawu biodrowego[7]. Niektórzy autorzy szacowali maksymalną prędkość allozaura na od 30 do 55 km/h[91], jednak inni spekulowali, że teropod ten mógł nie być w stanie poruszać się z prędkością przekraczającą 8 m/s (28,8 km/h)[7].
Zachowania stadne
Allosaurus jest od dawna opisywany w publikacjach popularnonaukowych oraz literaturze popularnej jako zwierzę polujące stadnie na zauropody i inną dużą zdobycz[12][26][65]. Robert Bakker rozszerzył zakres zachowań społecznych o opiekę rodzicielską – obecność zgubionych zębów i pogryzionych kości interpretował jako dowód na przynoszenie przez dorosłe osobniki jedzenia młodym oraz ochronę łupu przed innymi drapieżnikami[55]. Dowody na społeczne zachowania teropodów są jednak nikłe, a kontakty między osobnikami tego samego gatunku mogły obejmować również zachowania antagonistyczne, czego dowodzą ślady urazów na gastraliach[8], ślady ukąszeń na czaszkach (jak np. w przypadku patologicznie zniekształconej czaszce opisanej jako Labrosaurus ferox). Tego typu ugryzienia w głowę mogły służyć ustalaniu dominacji w stadzie lub rozwiązywaniu walk o terytorium[92].
Mimo iż allozaury mogły polować w stadach[93], nowsze badania sugerują, że teropody zamiast współpracy przejawiały zachowania agresywne wobec przedstawicieli tego samego gatunku. Naukowcy zauważyli, że wspólne polowanie na dużą ofiarę – często przypisywane teropodom – jest bardzo rzadkie u kręgowców w ogóle, również współczesne diapsydowe drapieżniki (m.in.: jaszczurki, krokodyle, ptaki) współpracują w ten sposób jedynie okazjonalnie. Zamiast tego są one typowo terytorialne oraz zabijają i zjadają przedstawicieli własnego gatunku. Zgodnie z tą interpretacją nagromadzenie szczątków allozaurów, jak np. w Cleveland-Lloyd, nie jest spowodowane zespołowym polowaniem, lecz faktem, że zwierzęta te gromadziły się, by żywić się martwymi lub słabymi allozaurami. Może to tłumaczyć wysoki odsetek młodych i młodocianych osobników na cmentarzyskach, gdyż są one znacznie częściej zabijane również na żerowiskach współczesnych drapieżników, takich jak krokodylowate i warany z Komodo. Interpretacja ta stoi w zgodności z teorią Bakkera[94]. Istnieją dowody na kanibalizm allozaurów, takie jak zęby odnalezione w pobliżu fragmentów żeber, ślady ugryzień na łopatkach[95] oraz szkielet allozaura wśród kości odnalezionych w miejscu uznanym przez Bakkera za gniazdo[81].
Mózg i zmysły
Mózg allozaura – według badań tomografowych puszki mózgowej – był bardziej podobny do mózgów krokodyli niż innych współczesnych archozaurów, ptaków. Struktura narządu statycznego dowodzi, że dinozaur ten trzymał czaszkę dokładnie poziomo. Ucho wewnętrzne budową przypominało krokodyle, więc Allosaurus mógł prawdopodobnie lepiej słyszeć niskie tony, a mieć problemy z wyższymi. Opuszki węchowe były duże i zdają się być dobrze wykształcone do wychwytywania zapachów, choć obszar służący do wychwytywania zapachów był stosunkowo niewielki[53].
Allozaur w kulturze masowej

Choć jest od niego dużo mniej znany i popularny, podobnie jak Tyrannosaurus allozaur stał się w kulturze masowej kwintesencją dużego mięsożernego dinozaura. Jest pospolitym dinozaurem w muzeach, głównie ze względu na wykopaliska w Cleveland-Lloyd Dinosaur Quarry. Od 1976 roku – w wyniku współpracy – 38 muzeów z ośmiu krajów na trzech kontynentach ma w swoich zbiorach skamieniałości pochodzące z Cleveland-Lloyd[5]. Allosaurus jest oficjalnym symbolem – skamieniałością stanu Utah (ang. state fossil – skamieniałość stanowa)[96].
Od początku XX wieku allozaur jest często przedstawiany w kulturze popularnej. To czołowy drapieżnik zarówno w powieści Arthura Conana Doyle’a Zaginiony świat z 1912, jak i jej filmowej adaptacji z 1925 – pierwszym pełnometrażowym filmie, w którym pojawiły się dinozaury[97]. Później pojawił się w filmie The Beast of Hollow Mountain z 1956[27] oraz w The Valley of Gwangi z 1969. Tytułowy Gwangi jest allozaurem, choć Ray Harryhausen jego postać wzorował na wizerunku tyranozaura z obrazu Charlesa R. Knighta[98]. Allozaur pojawił się także w filmie wytwórni Hammer Film Productions Milion lat przed naszą erą z 1966 oraz w The Land That Time Forgot z 1975. Spośród filmów popularnonaukowych allozaur był przedstawiony w drugim i piątym odcinku serialu BBC Wędrówki z dinozaurami, w Jurassic Fight Club, a także w specjalnym odcinku Wędrówek z dinozaurami – Balladzie o Wielkim Alu, który opowiada historię życia osobnika o przezwisku „Wielki Al”.
Przypisy
- 1 2 3 4 5 John R. Foster: Paleoecological Analysis of the Vertebrate Fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, U.S.A.. New Mexico Museum of Natural History and Science Bulletin 23: 37, 2003.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 Thomas R. Holtz Jr, Ralph E. Molnar, Philip J. Currie: Basal Tetanurae. W: David B. Weishampel, Peter Dodson, Halszka Osmólska (red.): The Dinosauria. Wyd. drugie. Berkeley: University of California Press, 2004, s. 71–110. ISBN 0-520-24209-2.
- 1 2 3 4 Allosaurus. W: Donald F. Glut: Dinosaurs: The Encyclopedia. Jefferson, North Carolina: McFarland & Co, 1997, s. 105–117. ISBN 0-89950-917-7.
- 1 2 3 4 5 Mickey Mortimer: And the Largest Theropod Is..... Dinosaur Mailing List. [dostęp 2008-09-28]. [zarchiwizowane z tego adresu (2010-03-25)]. (ang.).
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 James H. Madsen Jr: Allosaurus fragilis: A Revised Osteology. Wyd. drugie. Salt Lake City: Utah Geological Survey, 1993 [1976], seria: Utah Geological Survey Bulletin 109.
- 1 2 3 4 Allosaurus fragilis. W: John Foster: Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Bloomington, Indiana: Indiana University Press, 2007, s. 170–176. ISBN 978-0-253-34870-8. OCLC 77830875.
- 1 2 3 4 Karl T. Bates, Peter L. Falkingham, Brent H. Breithaupt, David Hodgetts, William I. Sellers, Phillip L. Manning. How big was 'Big Al'? Quantifying the effect of soft tissue and osteological unknowns of mass predictions for Allosaurus (Dinosauria: Theropoda). „Palaeontologia Electronica”. 12 (3), s. 14A, 2009. (ang.).
- 1 2 3 4 5 Daniel J. Chure: A new species of Allosaurus from the Morrison Formation of Dinosaur National Monument (Utah–Colorado) and a revision of the theropod family Allosauridae. Dysertacja doktorska. Columbia University, 2000.
- ↑ John Foster: Jurassic West: the Dinosaurs of the Morrison Formation and Their World. Bloomington, Indiana: Indiana University Press, 2007, s. 117.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 Genus Allosaurus. W: Gregory S. Paul: Predatory Dinosaurs of the World. New York: Simon & Schuster, 1988, s. 307–313. ISBN 0-671-61946-2.
- 1 2 Ralph E. Molnar. Analogies in the evolution of combat and display structures in ornithopods and ungulates. „Evolutionary Theory”. 3, s. 165–190, 1977. (ang.).
- 1 2 3 Carnosaurs. W: David B. Norman: The Illustrated Encyclopedia of Dinosaurs: An Original and Compelling Insight into Life in the Dinosaur Kingdom. New York: Crescent Books, 1985, s. 62–67. ISBN 0-517-468905.
- ↑ Gregory S. Paul: Predatory Dinosaurs of the World. New York: Simon & Schuster, 1988, s. 91, 93. ISBN 0-671-61946-2.
- ↑ Gregory S. Paul: Predatory Dinosaurs of the World. 1988, s. 277.
- ↑ Daniel J. Chure. Observations on the morphology and pathology of the gastral basket of Allosaurus, based on a new specimen from Dinosaur National Monument. „Oryctos”. 3, s. 39–37, 2000. ISSN 1290-4805. (ang.).
- ↑ Daniel J. Chure, James H. Madsen, On the presence of furculae in some non-maniraptoran theropods, „Journal of Vertebrate Paleontology”, 16 (3), 1996, s. 573–577, JSTOR: 4523744 (ang.).
- 1 2 3 4 5 6 Charles W. Gilmore. Osteology of the carnivorous dinosauria in the United States National Museum, with special reference to the genera Antrodemus (Allosaurus) and Ceratosaurus. „Bulletin of the United States National Museum”. 110, s. 1–159, 1920. (ang.).
- 1 2 3 Kenneth Carpenter. Forelimb biomechanics of nonavian theropod dinosaurs in predation. „Senckebergiana lethaea”. 82 (1), s. 59–75, 2002. DOI: 10.1007/BF03043773. (ang.).
- ↑ Genus Allosaurus. W: Gregory S. Paul: Predatory Dinosaurs of the World. New York: Simon & Schuster, 1988, s. 113. ISBN 0-671-61946-2.
- 1 2 Othniel Charles Marsh. Notice of new dinosaurian reptiles. „American Journal of Science and Arts”. 15, s. 241–244, 1878. (ang.).
- ↑ Friedrich von Huene. The carnivorous Saurischia in the Jura and Cretaceous formations, principally in Europe. „Revista del Museo de La Plata”. 29, s. 35–167, 1926.
- ↑ Alfred S. Romer: Osteology of the Reptiles. Chicago: University of Chicago Press, 1956. ISBN 0-89464985-X.
- ↑ Alfred S. Romer: Vertebrate Paleontology. Wyd. trzecie. Chicago: University of Chicago Press, 1966. ISBN 0-7167-1822-7.
- ↑ R. Steel: Part 14. Saurischia. Handbuch der Paläoherpetologie. Stuttgart: Gustav Fischer Verlag, 1970, s. 1–87.
- ↑ Alick D. Walker. Triassic reptiles from the Elgin area: Ornithosuchus and the origin of carnosaurs. „Philosophical Transactions of the Royal Society of London, Series B, Biological Sciences”. 248 (744), s. 53–134, 1964. DOI: 10.1098/rstb.1964.0009. (ang.).
- 1 2 Allosaurids. W: David Lambert, Diagram Group: The Dinosaur Data Book. New York: Avon Books, 1983, s. 80–81. ISBN 0-380-75896-2.
- 1 2 Allosaurids. W: David Lambert, Diagram Group: The Dinosaur Data Book. New York: Avon Books, 1990, s. 130. ISBN 0-380-75896-2.
- 1 2 Roger B. J. Benson, Matthew T. Carrano, Stephen L. Brusatte. A new clade of archaic large-bodied predatory dinosaurs (Theropoda: Allosauroidea) that survived to the latest Mesozoic. „Naturwissenschaften”. 97 (1), s. 71–78, 2010. DOI: 10.1007/s00114-009-0614-x. (ang.).
- ↑ The allosaur-tyrannosaur group. W: Gregory S. Paul: Predatory Dinosaurs of the World. 1988, s. 301–347.
- ↑ Thomas R. Holtz, Jr., The phylogenetic position of the Tyrannosauridae: Implications for theropod systematics, „Journal of Paleontology”, 68 (5), 1994, s. 1100–1117, JSTOR: 1306180 (ang.).
- ↑ Jerald D. Harris. A reanalysis of Acrocanthosaurus atokensis, its phylogenetic status, and paleobiological implications, based on a new specimen from Texas. „New Mexico Museum of Natural History and Science Bulletin”. 13, s. 1–75, 1998. (ang.).
- ↑ Paul C. Sereno, Didier B. Dutheil, M. Iarochene, Hans C. E. Larsson, Gabrielle H. Lyon, Paul M. Magwene, Christian A. Sidor, David J. Varricchio, Jeffrey A. Wilson. Predatory dinosaurs from the Sahara and Late Cretaceous faunal differentiation. „Science”. 272 (5264), s. 986–991, 1996. DOI: 10.1126/science.272.5264.986. PMID: 8662584. (ang.).
- ↑ Philip J. Currie, Kenneth Carpenter. A new specimen of Acrocanthosaurus atokensis (Theropoda, Dinosauria) from the Lower Cretaceous Antlers Formation (Lower Cretaceous, Aptian) of Oklahoma, USA. „Geodiversitas”. 22 (2), s. 207–246, 2000. (ang.).
- ↑ Joseph Leidy. Remarks on Poicilopleuron valens, Clidastes intermedius, Leiodon proriger, Baptemys wyomingensis, and Emys stevensonianus. „Proceedings of the Academy of Natural Sciences, Philadelphia”. 22, s. 3–4, 1870. (ang.).
- ↑ Joseph Leidy. Contribution to the extinct vertebrate fauna of the western territories. „Report of the U.S. Geological Survey of the Territories I”, s. 14–358, 1873. (ang.).
- ↑ Henry George Liddell, Robert Scott: Greek–English Lexicon, Abridged Edition. Oxford: Oxford University Press, 1980. ISBN 0-19-910207-4.
- 1 2 Othniel Charles Marsh. Notice of new dinosaurian reptiles from the Jurassic formation. „American Journal of Science and Arts”, s. 514–516, 1877. (ang.).
- ↑ Ben Creisler: Dinosauria Translation and Pronunciation Guide A. [dostęp 2009-02-12]. [zarchiwizowane z tego adresu (2011-11-06)]. (ang.).
- ↑ Othniel Charles Marsh. Principal characters of American Jurassic dinosaurs. Part II. „American Journal of Science, Series 3”. 17, s. 86–92, 1879. (ang.).
- ↑ Edward Drinker Cope. A new opisthocoelous dinosaur. „American Naturalist”. 12 (6), s. 406, 1878. DOI: 10.1086/272127. (ang.).
- ↑ Mark A. Norell, Eric S. Gaffney, Lowell Dingus: Discovering Dinosaurs in the American Museum of Natural History. New York: Knopf, 1995, s. 112–113. ISBN 0-679-43386-4.
- ↑ Brent H. Breithaupt, Daniel J. Chure, Elizabeth H. Southwell. AMNH 5753: The world's first free-standing theropod skeleton. „Journal of Vertebrate Paleontology”. 19 (3, Suppl.), s. 33A, 1999. DOI: 10.1080/02724634.1999.10011202. (ang.).
- ↑ Samuel Wendell Williston. American Jurassic dinosaurs. „Transactions of the Kansas Academy of Science”. 6, s. 42–46, 1878. (ang.).
- ↑ Samuel Wendell Williston. The dinosaurian genus Creosaurus, Marsh. „American Journal of Science, series 4”. 11 (11), s. 111–114, 1901. (ang.).
- ↑ Donald M. Henderson. Skull and tooth morphology as indicators of niche partitioning in sympatric Morrison Formation theropods. „Gaia”. 15, s. 219–266, 1998. ISSN 0871-5424. [zarchiwizowane z adresu 2011-07-19]. (ang.).
- ↑ William J. Stokes. A new quarry for Jurassic dinosaurs. „Science”. 101 (2614), s. 115–117, 1945. DOI: 10.1126/science.101.2614.115-a. PMID: 17799203. (ang.).
- ↑ Adrian P. Hunt, Spencer G. Lucas, Karl Krainer: The taphonomy of the Cleveland-Lloyd Dinosaur Quarry, Upper Jurassic Morrison Formation, Utah: a re-evaluation. W: John R. Foster, Spencer G. Lucas (red.): Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36: 57–65, 2006.
- ↑ Mark A. Loewen, Scott D. Sampson, Matthew T. Carrano, Daniel J. Chure. Morphology, taxonomy, and stratigraphy of Allosaurus from the Upper Jurassic Morrison Formation. „Journal of Vertebrate Paleontology”. 23 (3, Suppl.), s. 72A, 2003. DOI: 10.1080/02724634.2003.10010538. (ang.).
- 1 2 David K. Smith. A morphometric analysis of Allosaurus . „Journal of Vertebrate Paleontology”. 18 (1), s. 126–142, 1998. DOI: 10.1080/02724634.1998.10011039. (ang.).
- 1 2 Paul J. Bybee, A. H. Lee, E.-T. Lamm. Sizing the Jurassic theropod dinosaur Allosaurus: Assessing growth strategy and evolution of ontogenetic scaling of limbs. „Journal of Morphology”. 267 (3), s. 347–359, 2006. DOI: 10.1002/jmor.10406. (ang.).
- 1 2 John R. Foster, Daniel J. Chure: Hindlimb allometry in the Late Jurassic theropod dinosaur Allosaurus, with comments on its abundance and distribution. W: John R. Foster, Spencer G. Lucas (red.): Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36: 57–65, 2006.
- 1 2 Emily J. Rayfield, David B. Norman, Celeste C. Horner, John R. Horner i inni. Cranial design and function in a large theropod dinosaur. „Nature”. 409, s. 1033–1037, 2001. DOI: 10.1038/35059070. (ang.).
- 1 2 Scott W. Rogers, ''Allosaurus'', crocodiles, and birds: Evolutionary clues from spiral computed tomography of an endocast, „The Anatomical Record”, 257 (5), 1999, s. 163–173, DOI: 10.1002/(SICI)1097-0185(19991015)257:5<162::AID-AR5>3.0.CO;2-W (ang.).
- 1 2 Robert T. Bakker. Brontosaur killers: Late Jurassic allosaurids as sabre-tooth cat analogues. „Gaia”. 15, s. 145–158, 1998. ISSN 0871-5424. [zarchiwizowane z adresu 2011-07-19]. (ang.).
- 1 2 Robert T. Bakker: Raptor Family values: Allosaur parents brought giant carcasses into their lair to feed their young. W: Donald L. Wolberg, Edmund Sump, Gary D. Rosenberg (red.): Dinofest International, Proceedings of a Symposium Held at Arizona State University. Philadelphia: Academy of Natural Sciences, 1997, s. 51–63. ISBN 0-935868-94-1.
- ↑ Daniel J. Chure: A reassessment of the gigantic theropod Saurophagus maximus from the Morrison Formation (Upper Jurassic) of Oklahoma, USA. W: Ailing Sun, Yuangqing Wang (red.): Sixth Symposium on Mesozoic Terrestrial Ecosystems and Biota, Short Papers. Pekin: China Ocean Press, 1995, s. 103–106. ISBN 7-5027-3898-3.
- ↑ B.P. Pérez-Moreno, D.J. Chure, C. Pires, C.M. Silva i inni. On the presence of Allosaurus fragilis (Theropoda: Carnosauria) in the Upper Jurassic of Portugal: First evidence of an intercontinental dinosaur species. „Journal of the Geological Society”. 156 (3), s. 449–452, 1999. DOI: 10.1144/gsjgs.156.3.0449. (ang.).
- ↑ The discovery of a nearly complete Allosaurus from the Jurassic Morrison Formation, eastern Bighorn Basin, Wyoming. W: Brent Breithaupt, C.E. Brown, S. C. Kirkwood, T.S. Miller (red.): Forty-Seventh Annual Field Conference Guidebook. Casper, Wyoming: Wyoming Geological Association, 1996, s. 309–313. OCLC 36004754.
- 1 2 Howe Dinosaur Quarry. Bighorn Basin GeoScience Center. [dostęp 2011-08-06]. [zarchiwizowane z tego adresu (2017-10-12)]. (ang.).
- 1 2 Brent H. Breithaupt, The case of „Big Al” the ''Allosaurus'': a study in paleodetective partnerships, [w:] V.L. Santucci, L. McClelland (red.), Proceedings of the 6th Fossil Resource Conference, National Park Service, U.S. Department of the Interior, Geologic Resources Division Technical Report (NPS/NRGRD/GRDTR-01/01), 2001, s. 95–106 [zarchiwizowane z adresu 2011-10-18] (ang.).
- 1 2 3 Daniel J. Chure i Mark A. Loewen. Cranial anatomy of Allosaurus jimmadseni, a new species from the lower part of the Morrison Formation (Upper Jurassic) of Western North America. „PeerJ”. 8:e7803, 2020. DOI: 10.7717/peerj.7803. (ang.).
- 1 2 Rebecca R. Hanna, Multiple injury and infection in a sub-adult theropod dinosaur (Allosaurus fragilis) with comparisons to allosaur pathology in the Cleveland-Lloyd Dinosaur Quarry collection, „Journal of Vertebrate Paleontology”, 22 (1), 2002, s. 76–90, DOI: 10.1671/0272-4634(2002)022[0076:MIAIIA]2.0.CO;2 (ang.).
- ↑ Brent Breithaupt: The discovery of a nearly complete Allosaurus from the Jurassic Morrison Formation, eastern Bighorn Basin, Wyoming. W: C. E. Brown, S. C. Kirkwood, T. S. Miller: Forty-Seventh Annual Field Conference Guidebook. Casper, Wyoming: Wyoming Geological Association, 1996, s. 309–313.
- 1 2 Octávio Mateus, Aart Walen, Miguel Telles Antunes: The large theropod fauna of the Lourinha Formation (Portugal) and its similarity to that of the Morrison Formation, with a description of a new species of Allosaurus. W: John R. Foster, Spencer G. Lucas (red.): Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36: 123–129, 2006.
- 1 2 3 Allosaurus. W: Don Lessem, Donald F. Glut: The Dinosaur Society's Dinosaur Encyclopedia. Random House, 1993, s. 19–20. ISBN 0-679-41770-2. OCLC 30361459.
- ↑ David K. Smith. A discriminant analysis of Allosaurus population using quarries as the operational units. „Museum of Northern Arizona Bulletin”. 60, s. 69–72, 1996. (ang.).
- ↑ David K. Smith, Patterns of size-related variation within ''Allosaurus'', „Journal of Vertebrate Paleontology”, 19 (2), 1999, s. 402–403, JSTOR: 4524002 (ang.).
- ↑ Elisabete Malafaia, Pedro Dantas, Francisco Ortega, Fernando Escaso. Nuevos restos de Allosaurus fragilis (Theropoda: Carnosauria) del yacimiento de Andrés (Jurásico Superior; centro-oeste de Portugal). „Cantera Paleontológica”, s. 255–271, 2007. (hiszp.).
- ↑ E. Malafaia, F. Ortega, F. Escaso, P. Dantas, N. Pimentel, J.M. Gasulla, B. Ribeiro, F. Barriga, J. L. Sanz. Vertebrate fauna at the Allosaurus fossil-site of Andrés (Upper Jurassic), Pombal, Portugal. „Journal of Iberian Geology”. 36 (2), s. 193–204, 2010. DOI: 10.5209/rev_JIGE.2010.v36.n2.7. (ang.).
- ↑ Werner Janensch. Die Coelurosaurier und Theropoden der Tendaguru-Schichten Deutsch-Ostafrikas. „Palaeontographica, suplement 7”. 1, s. 1–99, 1925. (niem.).
- ↑ Oliver W. M. Rauhut. Post-cranial remains of 'coelurosaurs' (Dinosauria, Theropoda) from the Late Jurassic of Tanzania. „Geological Magazine”. 142 (1), s. 97–107, 2005. DOI: 10.1017/S0016756804000330. (ang.).
- ↑ Othniel Charles Marsh. Principal characters of American Jurassic dinosaurs. Part VIII. „American Journal of Science, Series 3”. 27, s. 329–340, 1884. (ang.).
- ↑ James H. Madsen, Samuel P. Welles: Ceratosaurus (Dinosauria, Theropoda), a Revised Osteology. Utah Geological Survey, 2000, seria: Miscellaneous Publication, 00-2.
- ↑ Ralph E. Molnar, Timothy F. Flannery, Thomas H. V. Rich. An allosaurid theropod dinosaur from the Early Cretaceous of Victoria, Australia. „Alcheringa”. 5, s. 141–146, 1981. DOI: 10.1080/03115518108565427. (ang.).
- ↑ Anatolij Nikołajewicz Riabinin. Zamjatka o dinozaury ise Zabajkalja. „Trudy Gieologicziesowo Muzjeja Imieni Pietra Wielkowo Impieratorskoj Akadiemii Nauk”. 8 (5), s. 133–140, 1914. (ros.).
- ↑ John R. Foster: Paleoecological Analysis of the Vertebrate Fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, U.S.A.. New Mexico Museum of Natural History and Science Bulletin 23: 29, 2003.
- ↑ Dale A. Russell: An Odyssey in Time: Dinosaurs of North America. Minocqua, Wisconsin: NorthWord Press, 1989, s. 175–176. ISBN 1-55971-038-1.
- ↑ Kenneth Carpenter: Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus. W: John R. Foster, Spencer G. Lucas (red.): Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36: 131–138, 2006.
- ↑ Daniel J. Chure, Ron Litwin, Stephen T. Hasiotis: The fauna and flora of the Morrison Formation: 2006. W: John R. Foster, Spencer G. Lucas (red.): Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36: 233–248, 2006.
- 1 2 Octávio Mateus: Jurassic dinosaurs from the Morrison Formation (USA), the Lourinhã and Alcobaça Formations (Portugal), and the Tendaguru Beds (Tanzania): A comparison. W: John R. Foster, Spencer G. Lucas (red.): Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36: 223–231, 2006.
- 1 2 Robert T. Bakker, Gary Bir: Dinosaur crime scene investigations: theropod behavior at Como Bluff, Wyoming, and the evolution of birdness. W: Philip J. Currie, Eva B. Koppelhus, Martin A. Shugar, Joanna L. Wright (red.): Feathered Dragons: Studies on the Transition from Dinosaurs to Birds. Bloomington i Indianapolis: Indiana University Press, 2004, s. 301–342. ISBN 0-253-34373-9.
- ↑ Daniel J. Chure, Anthony R. Fiorillo, Aase Jacobsen. Prey bone utilization by predatory dinosaurs in the Late Jurassic of North America, with comments on prey bone use by dinosaurs throughout the Mesozoic. „Gaia”. 15, s. 227–232, 2000. ISSN 0871-5424. [zarchiwizowane z adresu 2011-07-19]. (ang.).
- ↑ Andrew H. Lee, Sarah Werning. Sexual maturity in growing dinosaurs does not fit reptilian growth models. „Proceedings of the National Academy of Sciences”. 105 (2), s. 582–587, 2008. DOI: 10.1073/pnas.0708903105. PMID: 18195356. (ang.).
- ↑ Mark A. Loewen, Matthew T. Carrano, Scott D. Sampson. Ontogenetic changes in hindlimb musculature and function in the Late Jurassic theropod Allosaurus. „Journal of Vertebrate Paleontology”. 22 (3, Suppl.), s. 80A, 2002. (ang.).
- ↑ David E. Fastovsky: Dinosaur Paleoecology. W: David B. Weishampel, Joshua B. Smith, Peter Dodson, Halszka Osmólska (red.): The Dinosauria. Wyd. drugie. Berkeley: University of California Press, 2004, s. 614–626. ISBN 0-520-24209-2.
- ↑ Kenneth Carpenter, Frank Sanders, Lorrie A. McWhinney: Evidence for predator-prey relationships: Examples for Allosaurus and Stegosaurus. W: Kenneth Carpenter (red.): The Carnivorous Dinosaurs. Bloomington i Indianapolis: Indiana University Press, 2005, s. 325–350. ISBN 0-253-34539-1.
- ↑ T. H. Frazzetta, Kenneth V. Kardong. Prey attack by a large theropod dinosaur. „Nature”. 416, s. 387–388, 2002. DOI: 10.1038/416387a. (ang.).
- ↑ Emily J. Rayfield, David B. Norman, Paul Upchurch. Prey attack by a large theropod dinosaur: Response to Frazzetta and Kardong, 2002. „Nature”. 416, s. 388, 2002. DOI: 10.1038/416388a. (ang.).
- ↑ Kent A. Stevens, Binocular vision in theropod dinosaurs, „Journal of Vertebrate Paleontology”, 26 (2), 2006, s. 321–330, DOI: 10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2 (ang.).
- ↑ Evolve: Eyes. The History Channel. [dostęp 2009-03-27]. [zarchiwizowane z tego adresu (2008-12-11)]. (ang.).
- ↑ Per Christiansen. Strength indicator values of theropod long bones, with comments on limb proportions and cursorial potential. „Gaia”. 15, s. 241–255, 1998. (ang.).
- ↑ Darren H. Tanke, Philip J. Currie. Head-biting behavior in theropod dinosaurs: Paleopathological evidence. „Gaia”. 15, s. 167–184, 1998. ISSN 0871-5424. (ang.).
- ↑ Philip J. Currie: Theropods. W: James Farlow, Michael K. Brett-Surman (red.): The Complete Dinosaur. Indiana: Indiana University Press, 1999, s. 228. ISBN 0-253-21313-4. (ang.).
- ↑ Brian T. Roach, Daniel L. Brinkman, A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs, „Bulletin of the Peabody Museum of Natural History”, 48 (1), 2007, s. 103–138, DOI: 10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2 (ang.).
- ↑ Brandon Goodchild Drake. A new specimen of Allosaurus from north-central Wyoming. „Journal of Vertebrate Paleontology”. 24 (3, Suppl.), s. 65A, 2004. (ang.).
- ↑ Utah State Fossil – Allosaurus. Utah Department of Community and Culture. [dostęp 2009-03-28]. [zarchiwizowane z tego adresu (2016-03-03)]. (ang.).
- ↑ Donald F. Glut, Michael K. Brett-Surman: Dinosaurs and the media. W: James Farlow, Michael K. Brett-Surman (red.): The Complete Dinosaur. Bloomington i Indianapolis: Indiana University Press, 1997, s. 675–706. ISBN 0-253-33349-0.
- ↑ Return to the Valley, a documentary on the Region 1 Valley of Gwangi DVD.
- kategoria
- klasyfikacja
- lista
- wikiprojekt
| Infrarzędy dinozaurów | |
|---|---|
| Era dinozaurów | |
| Dyscypliny powiązane |
