| Triceratops | |||||
| Marsh, 1889[1] | |||||
| Okres istnienia: Kreda późna, 68–65 mln lat temu | |||||
 Prawie dorosły T. horridus | |||||
| Systematyka | |||||
| Domena | |||||
|---|---|---|---|---|---|
| Królestwo | |||||
| Typ | |||||
| Podtyp | |||||
| Gromada | |||||
| Podgromada | |||||
| Nadrząd | |||||
| Rząd | |||||
| Podrząd | |||||
| Infrarząd | |||||
| Rodzina | |||||
| Podrodzina | |||||
| Plemię |
Triceratopsini | ||||
| Rodzaj |
triceratops | ||||
| |||||
| Gatunki | |||||
| |||||
Triceratops[uwaga 1] – rodzaj roślinożernego dinozaura rogatego z rodziny ceratopsów, żyjącego w mastrychcie (kreda późna), od 68 do 65,5 milionów lat temu na terenach dzisiejszej Ameryki Północnej. Należy do ostatnich nieptasich dinozaurów, które pojawiły się przed wymieraniem kredowym. Nazwa rodzajowa pochodzi z greki i oznacza twarz o trzech rogach.
Nosząc dużą kostną kryzę na szyi i trzy rogi, czworonożny, przypominający pod pewnymi względami nosorożca triceratops stał się jednym z najbardziej rozpoznawalnych dinozaurów i najlepiej znanym przedstawicielem Ceratopsidae. Dzielił środowisko życia i stanowił zdobycz tyranozaura[2], choć nie jest pewne, czy te dwa dinozaury rzeczywiście walczyły ze sobą w sposób, w jaki się to często przedstawia na popularnych obrazkach.
Dokładne umiejscowienie rodzaju wśród ceratopsydów stanowi przedmiot debaty paleontologów. Obecnie uznaje się dwa pewne gatunki: T. horridus i T. prorsus, chociaż nazywano liczne gatunki. Badanie opublikowane w 2012 sugeruje też, że współczesny mu torozaur, marginocefal długo uznawany za odrębny rodzaj, stanowi w rzeczywistości dojrzałą formę triceratopsa[3][4]. Nie zostało to jednak zaakceptowane przez wszystkich specjalistów[5]. Publikacja Daniela Fielda i Nicholasa Longricha, badaczy z Yale, z 2012 sprzeciwiła się temu stwierdzeniu, pozostając przy odrębnej klasyfikacji jako oddzielne gatunki[6][7].
Znaleziono liczne szczątki triceratopsa, odkąd po raz pierwszy opisano rodzaj w 1889. Wlicza się w to co najmniej jeden kompletny szkielet. Na obszarze formacji Hell Creek tylko w pierwszej dekadzie XXI wieku znaleziono 47 kompletnych bądź fragmentarycznych czaszek. Odkryto osobniki reprezentujące różne stadia życia od wylęgających się młodych po dorosłe.
Funkcja kryzy kostnej i trzech wydatnych rogów twarzowych długo inspirowały do debaty. Tradycyjny pogląd widzi je jako broń służącą do obrony przed drapieżnikami. Bardziej współczesne pomysły zauważają obecność naczyń krwionośnych na kościach czaszki Ceratopsidae, uznając za bardziej prawdopodobne, że cechy te służyły identyfikacji, parowaniu się i wykazywaniu dominacji w grupie, jak poroża czy rogi dzisiejszych kopytnych czy chrząszczów. Teoria ta znalazłaby dodatkowe wsparcie, gdyby torozaur okazał się dojrzałą formą triceratopsa. Oznaczałoby to, że charakterystyczne dla torozaurów okna w kryzie pojawiały się dopiero z osiągnięciem dojrzałości przez osobniki, czyniąc kryzę bardziej przydatną do pokazywania niż do obrony.
Budowa
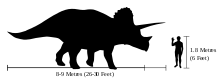
Długość triceratopsa szacuje się na około 7,9–9,0 m, wysokość na 2,9–3,0 m[8][9], masę ciała zaś na 6,1–12,0 ton[10]. Najbardziej wyróżnia się on swą wielką czaszką, należącą do najbardziej okazałych wśród zwierząt lądowych. Największa znana czaszka, okaz BYU 12183, miałaby całkowitą długość szacowaną na 2,5 m, gdyby była kompletna[3]. Mogło to stanowić prawie jedną trzecią długości całego zwierzęcia[11]. Na pysku, nad nozdrzami, sterczał pojedynczy róg, natomiast para rogów mierzących około metra długości znajdowała się nad oczami. Ze sklepienia czaszki odchodziła względnie krótka kryza kostna, zdobiona kośćmi epokcypitalnymi u niektórych okazów. Większość innych Ceratopsidae miała w kryzie duże okna, podczas gdy u triceratopsa tworzyła ją lita kość.
Skóra triceratopsa wyróżniała go wśród dinozaurów. Odciski skóry nieopisanego jeszcze osobnika ukazują, że niektóre gatunki mogły być pokryte przypominającymi włosy tworami, podobnymi do znalezionych u prymitywnego ceratopsa psitakozaura[12].
Kończyny
Triceratops cechował się potężną budową. Miał silne kończyny o krótkich dłoniach zakończonych trzema kopytkami każda i także krótkich stopach po cztery kopytka[13]. Choć pewnie był czworonożny, postawa triceratopsa pozostawała długo przedmiotem dyskusji. Pierwotnie sądzono, że przednie nogi gada były rozcapierzone na boki pod kątem w stosunku do tułowia, by lepiej nosić ciężar głowy[14]. Ilustrują to rysunki Charlesa Knighta i Rudolpha Zallingera. Dowody ichnologiczne w postaci skamieniałych śladów dinozaurów rogatych i współczesne rekonstrukcje szkieletów (i fizyczne, i cyfrowe) zdają się wykazywać, że triceratops i inne ceratopsydy utrzymywały pozycję wyprostowaną podczas zwyczajnej lokomocji, z łokciami zgiętymi i lekko pochylonymi, w pozycji pośredniej pomiędzy pełnym wyprostem i rozcapierzeniem, jak u współczesnego nosorożca[13][15][16][17].
Przedramiona i dystalne od nich części kończyny górnej triceratopsa zachowały prymitywną budowę w porównaniu z innymi czworonożnymi dinozaurami, jak tyreofory czy zauropody. W tych dwóch grupach przedramiona gatunków czworonożnych były zazwyczaj zrotowane tak, że ręce zwracały się ku przodowi z dłońmi odchylonymi w tył w pronacji, gdy zwierzę chodziło. Triceratops, jak inne ceratopsy i blisko spokrewnione ornitopody, chodził z większością palców skierowaną na zewnątrz, od ciała. Ten prymitywny jak na dinozaura stan spotyka się również u form dwunożnych, np. teropodów. U triceratopsa masa ciała spoczywała jedynie na pierwszych trzech palcach ręki. Trzeci i czwarty zachowały się w stanie szczątkowym, nie miały pazurów ni kopytek[13]. Wzór paliczków to 2-3-4-3-1, co oznacza, że najbardziej wewnętrzny palec składał się z dwóch kości, kolejny z trzech i tak dalej[18].
Systematyka

Nazwa rodzajowa Triceratops oznacza „trójrogą twarz”. Pochodzi ze starożytnej greki od τρί- (tri-, czyli „trzy”), κέρας (kéras, „róg”) i ὤψ (ops, „twarz”)[19][20].
Triceratops stanowi najlepiej poznany rodzaj z rodziny ceratopsów (Ceratopsidae, ceratopsydy) obejmującej duże ceratopsy (Ceratopsia, dinozaury rogate) z Ameryki Północnej. Dokładne miejsce triceratopsa wśród Ceratopsidae podlegało długiej debacie. Niepewność ta wywodzi się głównie z kombinacji krótkiej, jednolitej kryzy kostnej, przypominającej kryzy centrozaurów, i długich rogów brwiowych, podobnych do występujących u chasmozaurów. W pierwszym przeglądzie dinozaurów rogatych Richard Swann Lull zaproponował hipotezę istnienia dwóch linii. Pierwsza zawierała monokloniusa i centrozaura i właśnie ona prowadziła do triceratopsa. Kolejna obejmowała ceratopsa i torozaura. Czyniłoby to triceratopsa centrozaurynem w dzisiejszym rozumieniu tego słowa[21]. Późniejsze rewizje wsparły ten pogląd, formalnie opisując drugą grupę o długich kryzach jako chasmozauryny (Chasmosaurinae)[22][23].
Jako pierwszy zakwestionował powyższy pogląd w 1949 Charles Mortram Sternberg, uznając, że triceratops był bliżej spokrewniony z Arrhinoceratops i chasmozaurem. Uczony bazował na cechach czaszek i rogów. Włączył więc triceratopsa w Ceratopsinae, które określał jako Chasmosaurinae[24]. W większości go zignorowano. Zarówno John Ostrom[25], jak i później David Norman umieszczali triceratopsa w Centrosaurinae[26].
Kolejne odkrycia i analizy podtrzymały jednak pogląd Sternberga na pozycje systematyczną „trójrogiej twarzy”. Lehman zdefiniował obie podrodziny, uznając triceratopsa za chasmozauryna, opierając się na kilku cechach morfologicznych. W rzeczywistości pasował on dobrze do podrodziny Ceratopsinae, nie licząc jednej cechy: skróconej kryzy[27]. Późniejsze badania wykonane przez Petera Dodsona, włączając w to analizę kladystyczną z 1990[28] i badanie z 1993 z użyciem RFTRA (resistant-fit theta-rho analysis)[29], techniki morfometrycznej systematycznie mierzącej podobieństwa w kształcie czaszek, wzmocniły tezę o przynależności triceratopsa do ceratopsynów.
Kladogram z pracy Sampson et al. (2010)[30]
| Ceratopsidae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Użycie w filogenetyce
W taksonomii filogenetycznej triceratopsa wykorzystano jako punkt odniesienia przy tworzeniu definicji dinozaurów. Dinosauria określono jako wszystkich potomków ostatniego wspólnego przodka triceratopsa i ptaków nowoczesnych Neornithes[31]. Idąc dalej, dinozaury ptasiomiedniczne określono jako wszystkie dinozaury, których ostatni wspólny przodek z triceratopsem pojawił się później, niż ostatni wspólny przodek ich i ptaków nowoczesnych Neornithes[32].
Pochodzenie i ewolucja

Zwierzę żyło w mastrychcie (kreda późna), od 68 do 65,5 milionów lat temu, na terenach dzisiejszej Ameryki Północnej. Należało do ostatnich nieptasich dinozaurów, które pojawiły się przed wymieraniem kredowym[33].
Przez wiele lat po odkryciu ewolucyjne początki triceratopsa pozostawały w większości ukryte. W 1922 nowo odkryty protoceratops wydawał się jego przodkiem Osbornowi[34], ale minęło wiele dziesiątków lat, nim kolejne odkrycia ujrzały światło dzienne. Ostatnie lata okazały się bogate w okrycia dinozaurów spokrewnionych z przodkiem „trójrogiej twarzy”. Zuniceratops, najwcześniejszy znany dinozaur rogaty o rogach brwiowych, opisany został w późnych latach dziewięćdziesiątych XX wieku, a pierwszy znany dinozaur rogaty w ogóle, jurajski jinlong, w 2005.
Te nowe znaleziska okazały się istotne w ilustrowaniu początków dinozaurów rogatych w ogólności, sugerując ich azjatyckie pochodzenie z jury i rozprzestrzenienie się rzeczywiście rogatych dinozaurów na początku kredy późnej w Ameryce Północnej[35]. Jako że triceratops jest coraz pewniej uznawany za członka podrodziny długokryzych chasmozaurynów, jego przodek prawdopodobnie mógł przypominać chasmozaura, panującego 5 milionów lat wcześniej.
Paleobiologia

Choć triceratopsa przedstawia się zazwyczaj jako zwierzę stadne, istnieją właściwie niewielkie dowody na stadny tryb życia tego zwierzęcia. Podczas gdy kilka innych rodzajów dinozaurów rogatych znanych jest z cmentarzysk zawierających kości od dwóch do setek, a nawet tysięcy osobników, dotychczas udokumentowano jedno cmentarzysko zdominowane przez kości triceratopsa. Miejsce to leży na południowym wschodzie Montany, obejmuje szczątki trzech osobników młodocianych. Obecność tylko triceratopsów w tym wieku zwraca jednak uwagę[36]. Inne, bardziej współczesne znalezisko może ujawnić, że triceratopsy żyły w niewielkich grupach rodzinnych. W 2012 grupę trzech osobników, względnie kompletnych, znaleziono w Wyoming, w okolicy Newcastle. Każdy z nich był innej wielkości: od w pełni wyrośniętego do małego. Szczątki są obecnie wydobywane przez zespół paleontologów pod kierunkiem Petera Larsona z Black Hills Institute. Uważa się, że zwierzęta podróżowały jako grupa rodzinna, ale pozostaje tajemnicą, czy grupa składała się z pary i ich potomka, czy dwóch samic opiekujących się osobnikiem młodocianym. Pozostałości tego ceratopsa wykazują także ślady drapieżnictwa bądź padlinożerności tyranozaura, zwłaszcza największy osobnik. Kości przedniej kończyny tego ptasiomiedniczego uległy złamaniu i ranom penetrującym wywołanym przez zęby tyranozaura[37].
Przez wiele lat znajdywano jedynie szczątki samotnych osobników triceratopsa[36]. Są one bardzo pospolite. Na przykład paleontolog Bruce Erickson, pracujący dla Science Museum of Minnesota, odnotował, że widział 200 okazów T. prorsus w montańskiej formacji Hell Creek[38]. Podobnie Barnum Brown twierdził, że widział ponad 500 czaszek w terenie[39]. Ponieważ triceratopsie zęby, kawałki rogów i kryzy oraz inne fragmenty czaszki należą do skamielin znajdywanych obficie w faunie Lancian z późnego mastrychtu (kreda późna, od 68 do 65 milionów lat temu) na zachodzie dzisiejszej Ameryki Północnej, zwierzę uznaje się za dominującego roślinożercę tych czasów. W 1986 Robert Bakker oszacował, że przypadało nań do 5/6 fauny dużych dinozaurów pod koniec okresu kredowego[40]. W przeciwieństwie do większości zwierząt, skamieniałości czaszek są znacznie częstsze od pozostałości szkieletu pozaczaszkowego triceratopsa, co wskazuje na niezwykle duży potencjał zachowywania się czaszek[41].
Triceratops należał do ostatnich ceratopsów, jakie pojawiły się przed wymieraniem kredowym. Występowały wtedy także blisko spokrewniony z nim torozaur i dalszy krewny, niewielki leptoceratops, aczkolwiek ich szczątki są znacznie rzadsze[14].
Uzębienie i dieta

Triceratops był roślinożercą. Trzymał swoją głowę nisko, więc prawdopodobnie żywił się nisko rosnącą roślinnością, choć być może potrafił powalać wyższe rośliny rogami, dziobem i masą ciała[35][42]. Żuchwa i szczęka kończyły się głębokim, wąskim dziobem, prawdopodobnie lepiej przystosowanym do chwytania i zrywania, niż do gryzienia[25].
Zęby triceratopsa tworzyły grupy zwane bateriami zębowymi. Występowało od 36 do 40 kolumn zębów po każdej stronie szczęki czy żuchwy, w każdej z nich leżało od 3 do 5 ułożonych na stos zębów. Ta ostatnia wartość zależała od wielkości zwierzęcia[35]. Daje to w efekcie zakres od 432 do 800 zębów, z których jedynie ułamek służył zauropsydowi kiedykolwiek, zastępowanie zębów było procesem ciągłym i zachodziło przez całe życie zwierzęcia[35]. Pracowały one, tnąc w kierunku pionowym bądź zbliżonym do niego[35]. Wielki rozmiar i liczne zęby triceratopsa sugerują, że zjadał on wielkie ilości włóknistego pokarmu roślinnego[35]. Niektórzy widzą tutaj palmowce i sagowcowate[43][44], a inni wskazują na paprocie porastające wtedy tereny zajmowane przez współczesne prerie[45].
Funkcje rogów i kryzy
Wiele spekulacji dotyczyło funkcji pełnionych przez struktury zdobiące triceratopsią głowę. Powstały dwie główne teorie, wskazujące na użycie ich w walkach bądź do pokazów w celu zdobycia partnera. Obecnie ta druga wydaje się główną funkcją[35].
Na wczesnym etapie badań Lull postulował, że kryza stanowiła miejsce przyczepu dla mięśni szczęk, umożliwiających żucie. Pozwoliłoby to na większe rozmiary tychże mięśni, a więc umożliwiałoby im to rozwijanie większej siły[46]. Inni autorzy uznawali ten pogląd przez lata, ale późniejsze studia nie wykazały żadnych pozostałości po przyczepach mięśni na kościach tworzących kryzę[47].
Długo sądzono, że Triceratops prawdopodobnie wykorzystywał swoje rogi i kryzę w walkach z drapieżnikami takimi, jak tyranozaur. Pomysł ten przedyskutował jako pierwszy C. H. Sternberg w 1917, a 70 lat po nim zrobił to Robert Bakker[48][49]. Wykazano, że tyranozaur rzeczywiście miewał bezpośrednie pełne agresji spotkania z triceratopsem, opierając się na częściowo wygojonych śladach tyranozaurzych zębów na rogach brwiowych triceratopsa i jego kościach łuskowych. Ugryziony róg jest również złamany, po złamaniu wytworzyła się nowa kość. Nie wiadomo jednak, które zwierzę było agresorem[50]. Ponieważ rany triceratopsa zagoiły się, wydaje się bardzo prawdopodobne, że przeżył on to spotkanie i starał się pokonać teropoda. Paleontolog Peter Dodson spekuluje, że w przypadku zaatakowania przez tyranozaura potężnego triceratopsa ten ostatni miał przewagę i mógł z powodzeniem obronić się, zadając drapieżnikowi śmiertelne rany, używając swych ostrych rogów[51]. Wiadomo też, że tyranozaur żywił się triceratopsami. Dowodzą tego ciężko pogryzione triceratopsie kość biodrowa i kość krzyżowa[2].

Poza walkami z drapieżnikami z użyciem swych rogów triceratopsy przedstawia się tradycyjnie walczące między sobą z zablokowanymi rogami. Badania wykazały, że takie zachowanie byłoby możliwe, jeśli różniło się od poczynań dzisiejszych zwierząt rogatych[52], jednak nie ma zgody, czy rzeczywiście występowało. Chociaż znaleziono jamki, dziury, zmiany i inne uszkodzenia triceratopsich czaszek (jak też czaszek innych ceratopsów), które często przypisuje się obrażeniom odniesionym w takich walkach, współczesne badanie nie znajduje dowodów, że urazy te zostały zadane rogiem. Nie ma choćby śladów infekcji ani gojenia. Zamiast tego zasugerowano inne wyjaśnienia: niepatologiczną resorpcję kości czy też nieznaną ich chorobę[53]. Nowsza praca porównuje zapadalność zmian czaszkowych i reakcję okostnową triceratopsa i centrozaura. Wykazała ona ich zgodność u tego pierwszego, używającego swych rogów w walce i cechującego się kryzą przystosowaną do ochrony. Natomiast niższa zapadalność u centrozaura może wskazywać na to, że omawiane struktury jego głowy pełniły bardziej funkcję dekoracyjną lub też, że wykorzystywał on w zmaganiach inne części ciała zamiast głowy[54]. Częstość urazu u tricersatopsa wynosi 14%[55]. Badacze zauważyli również, że uszkodzenia znalezione na zbadanych okazach miały zbyt zlokalizowany charakter, by wynikać z choroby kości[56]. Badanie histologiczne ujawniło, że kryzę triceratopsa buduje kość o budowie włóknisto-blaszkowatej[57], zawierająca fibroblasty pełniące kluczową rolę w gojeniu się ran, zdolne do szybkiego odkładania kości w trakcie remodelingu[58][59].

Bardziej współczesne pomysły zauważają obecność naczyń krwionośnych na kościach czaszki Ceratopsidae, uznając za bardziej prawdopodobne, że cechy te służyły identyfikacji, parowaniu się i wykazywaniu dominacji w grupie, jak poroża czy rogi dzisiejszych reniferów, kozłów śnieżnych czy chrząszczów rohatyńcowatych[14]. Teoria ta znalazłaby dodatkowe wsparcie, gdyby torozaur okazał się dojrzałą formą triceratopsa. Jego kryza rozwijałaby okna z rozwinięciem dojrzałości, co wiązało się z przydatnością kryzy bardziej do pokazywania niż do obrony[3].
Duża kryza mogła także pomagać w termoregulacji, zwiększając powierzchnię ciała[60]. Podobny pomysł zaproponowano w przypadku płyt stegozaura[61], aczkolwiek taka rola nie wyjaśnia dziwacznych i wyszukanych wariacji spotykanych u różnych członków rodziny ceratopsów[35]. Jest to bardzo sugestywna obserwacja, wobec czego dzisiaj uważa się, że ornamenty te rosły głównie na pokaz.
Teoria upatrująca roli tych struktur w pokazach godowych zaproponowana została po raz pierwszy przez Davitashviliego w 1961. Zdobyła od tego czasu szeroką akceptację[27][47][62]. Istnieją przesłanki za ważną rolą pokazów, zarówno w doborze partnera, jak i w innych zachowaniach społecznych. Wskazuje na to znaczne zróżnicowanie ozdób dinozaurów rogatych, używanych w podobny sposób[63]. Niedawne badanie najmniejszej z czaszek triceratopsa, przypisywanej osobnikowi młodocianemu, wykazuje obecność kryzy i rogów rozwiniętych już na bardzo wczesnym etapie rozwoju, jeszcze przed dojrzewaniem płciowym, co wskazuje na ważną rolę komunikacji wzrokowej i rozpoznawanie się gatunków w ogólności[64].
Paleopatologia
W jednej z przypisywanych triceratopsowi czaszek dostrzeżono otwór w kości jarzmowej. Zdawał się on stanowić ranę penetrującą, którą zwierzę otrzymało za życia. Tezę tą wspierają pozostałości gojenia zachowane na kości wokół podejrzewanej rany. Dokładne badanie ujawnia, że średnica rany odpowiada średnicy dystalnego końca rogu triceratopsa. Tę i inne widoczne wygojone rany na czaszkach ceratopsów podawano jako dowód na nieprowadzącą do śmierci konkurencję wewnątrzgatunkową tej grupy dinozaurów[65][66].
Odkrycie

Pierwszy nazwany okaz przypisywany obecnie triceratopsowi to para rogów brwiowych umocowanych na sklepieniu czaszki, znalezionych w okolicy Denver wiosną 1887[67]. Okaz wysłano Otnielowi C. Marshowi, który sądził, że formacja, z której pochodzi, powstała z pliocenu. Uznał kości za należące do szczególnie dużego i niezwykłego bizona, którego nazwał Bison alticornis[67][68]. Następnego roku zorientował się, że były tam rogate dinozaury. Wtedy też opisał rodzaj Ceratops, bazując na fragmentarycznych szczątkach[69], cały czas uważał jednak B. alticornis za plioceńskiego ssaka. Dopiero dzięki trzeciej, znacznie bardziej kompletnej czaszce zmienił zdanie. Okaz zebrany w 1888 przez Johna B. Hatchera w formacji Lance na terenie Wyoming został pierwotnie opisany jako inny gatunek rodzaju Ceratops[70]. Po namyśle Marsh zmienił poglądy i nadał znalezisku nazwę rodzajową Triceratops, uznając Bison alticornis za inny gatunek Ceratops[71] (później przeniesiono go do Triceratops[21]). Solidna budowa czaszki zwierzęcia przyczyniła się do zachowania wielu okazów jako skamieniałości, co pozwoliło na przestudiowanie różnic międzygatunkowych i międzyosobniczych. Pozostałości triceratopsa znajdowano następnie w amerykańskich stanach Montana i Dakota Południowa (poza Colorado i Wyoming), a także w kanadyjskich prowincjach Saskatchewan i Alberta.
Wcześniejszy okaz, także odnaleziony w formacji Lance, nazwany został Agathaumas sylvestris przez Edwarda D. Cope’a w 1872. Pierwotnie zidentyfikowany jako kaczodzioby, składał się tylko z pozostałości szkieletu pozaczaszkowego i tylko tymczasowo uznaje się go za okaz triceratopsa[72].
Od tamtych czasów znaleziono liczne szczątki triceratopsa, odkąd po raz pierwszy opisano rodzaj w 1889. Wlicza się w to co najmniej jeden kompletny szkielet[13]. Paleontolog John Scannella stwierdził nawet, że trudno jest chodzić po formacji Hell Creek i nie potknąć się o triceratopsa z wietrzejącego zbocza. W tym obszarze tylko w pierwszej dekadzie XXI wieku znaleziono 47 kompletnych bądź fragmentarycznych czaszek[73]. Odkryto osobniki reprezentujące różne stadia życia od wylęgających się młodych po dorosłe[11].
Gatunki
Przez pierwsze dekady po opisaniu triceratopsa zbierano różne czaszki, różniące się w mniejszym bądź większym stopniu od oryginalnego „trójrogiego oblicza”, nazwanego T. horridus przez Marsha. Łaciński przymiotnik horridus oznacza „szorstki, chropowaty”. Epitet gatunkowy odnosi się do takiej tekstury kości należących do holotypu, później zidentyfikowanego jako osobnik w starszym wieku. Różnice w znajdowanych czaszkach nie zaskakują. Chodzi o duże, trójwymiarowe obiekty pochodzące od zwierząt różnego wieku i płci, poddane różnym ciśnieniom działającym z różnych stron w procesie fosylizacji[14]. Odkrywcy nadawali im miana oddzielnych gatunków i tworzyli różne schematy filogenetyczne przedstawiające, jak miały być ze sobą spokrewnione poszczególne gatunki.

Podejmując pierwszą próbę zrozumienia bogactwa gatunków, Lull znalazł dwie grupy, aczkolwiek nie zdradził, w jaki sposób je rozróżnił. Pierwsza składała się z T. horridus, T. prorsus i T. brevicornus; druga z T. elatus i T. calicornis. 2 gatunki (T. serratus i T. flabellatus) pozostały poza grupami[21]. W 1933 w jego rewizji kluczowej monografii z 1907 autorstwa Hatcher-Marsh-Lull wszystkich znanych ceratopsów pozostawił dwie grupy i dwa nieprzypisane nigdzie gatunki, oprócz nich uznając trzecią linię z T. obtusus i T. hatcheri, charakteryzującą się bardzo małym rogiem nosowym[23]. C. M. Sternberg wprowadził jedną modyfikację, dodając T. eurycephalus i sugerując, że leżał on bliżej drugiej i trzeciej linii, niż gałąź T. horridus[24]. Układ ten popierano aż do większych badań w latach osiemdziesiątych i dziewięćdziesiątych XX wieku.
Z czasem pomysł, zgodnie z którym różniące się od siebie czaszki mogą reprezentować nie oddzielne gatunki, a różnice międzyosobnicze w obrębie jednego-dwóch gatunków, zdobył sobie popularność. W 1986 Ostrom i Wellnhofer opublikowali pracę, gdzie zaproponowali istnienie pojedynczego gatunku Triceratops horridus[74]. Oparli się o przesłankę istnienia tylko jednego-dwóch gatunków dużych ssaków w danym regionie. Współczesny przykład stanowić miał słoń afrykański i żyrafa w dzisiejszej Afryce (obecnie wyróżniany jest jeszcze drugi gatunek słonia, słoń leśny[75]). Do ich wniosków Lehman dodał stare linie Lull-Sternberg w połączeniu z różnicami wiekowymi i dymorfizmem płciowym, sugerując, że linia T. horridus-T. prorsus-T. brevicornus składała się z samic, natomiast T.calicornis-T.elatus były samcami, a gałąź T. obtusus-T. hatcheri to z kolei stare samce, cierpiące z powodu patologii[27]. Zgodnie z jego rozumowaniem samce miały dłuższe, bardziej wyprostowane rogi i większe czaszki, samice zaś miały mniejsze czaszki o krótszych, skierowanych do przodu rogach.

Wnioski te zakwestionowała kilka lat później Catherine Forster. Powtórnie przeanalizowała ona materiał triceratopsa w bardziej wyczerpujący sposób. Doszła do wniosku, że szczątki należą do dwóch gatunków: T. horridus i T. prorsus, choć wyróżniająca się czaszka T. („Nedoceratops”) hatcheri była do tego stopnia inna, by wydzielić ją do osobnego rodzaju[76]. Uczona pokazała, że T. horridus i kilka innych „gatunków” leżało razem, a T. prorsus i T. brevicornus stało samotnie. Ponieważ w tej pierwszej grupie było znacznie więcej okazów, badaczka zasugerowała, że te dwie grupy reprezentują odrębne gatunki. Cały czas pozostaje możliwość interpretacji wyróżniającej pojedynczy gatunek wykazujący dymorfizm płciowy[14][77].
W 2009 John Scannella i Denver Fowler wsparli rozdzielenie T. prorsus i T. horridus. Zauważyli, że te dwa gatunki rozdzielone są także stratygraficznie w formacji Hell Creek. Oznacza to, że nie mogły żyć razem w tym samym czasie[78].
Pewne gatunki
- T. horridus (Marsh, 1889) (pierwotnie Ceratops) (gatunek typowy)
- T. prorsus (Marsh, 1890)
Synonimy i gatunki niepewne


Wymienione niżej gatunki uznaje się za nomina dubia (co oznacza „nazwy wątpliwe”). Bazują one na szczątkach zbyt ubogich, niekompletnych, by odróżnić je od uprzednich gatunków triceratopsa
- T. albertensis (C. M. Sternberg, 1949)
- T. alticornis (Marsh, 1887 [pierwotnie Bison])
- T. brevicornus (Hatcher, 1905) (=T. prorsus)
- T. calicornis (Marsh, 1898) (=T. horridus)
- T. elatus (Marsh, 1891) (=T. horridus)
- T. eurycephalus (Schlaikjer, 1935)
- T. flabellatus (Marsh, 1889) (=T. horridus)
- T. galeus (Marsh, 1889)
- T. hatcheri (Lull, 1907) (kontrowersyjny; zob. niżej Nedoceratops)
- T. ingens (Lull, 1915)
- T. maximus (Brown, 1933)
- T. mortuarius (Cope, 1874) (nomen dubium; pierwotnie Polyonax mortuarius)
- T. obtusus (Marsh, 1898) (=T. horridus)
- T. serratus (Marsh, 1890) (=T. horridus)
- T. sulcatus (Marsh, 1890)
- T. sylvestris (Cope, 1872) (nomen dubium; pierwotnie Agathaumas sylvestris)
Nedoceratops

Publikacja opisująca nedoceratopsa stanowiła pierwotnie część opus magnum Marsha, jego monografii Ceratopsidae. Niestety paleontolog zmarł w 1899, nie ukończywszy pracy. John Bell Hatcher podjął próbę ukończenia części poświęconej triceratopsowi. Zmarł jednak na tyfus plamisty w 1904, mając zaledwie 42 lata, pozostawiając publikację nieukończoną. Dokończyć dzieło przypadło w 1905 Richardowi S. Lullowi. Opublikował on wykonany przez Hatchera opis czaszki oddzielnie, nadając znalezisku nazwę Diceratops hatcheri[79][80]. Diceratops znaczy „dwuroga twarz”.
Ponieważ opis Diceratops stworzony został przez Hatchera, a Lull jedynie nadał okazowi nazwę i opublikował pracę po śmierci autora, Lull nie był zbytnio przekonany o odrębności Diceratops, uznając różnice za przejaw patologii. W 1933 Lull przemyślał ponownie uznawanie Diceratops za odrębny rodzaj. Uznał go za podrodzaj triceratopsa: Triceratops (Diceratops), włączając T. obtusus; istniejące różnice w większości przypisał podeszłemu wiekowi zwierzęcia.
Ponieważ nazwa Diceratops została wcześniej użyta dla owada błonkoskrzydłego (Foerster, 1868), Andriej Siergiejewicz Ukrainski nadał zwierzęciu jego obecną nazwę Nedoceratops w 2007[81]. Nieświadom tego, Octávio Mateus stworzył inną nową nazwę w 2008: Diceratus[82][83]. Diceratus stał się więc młodszym synonimem Nedoceratops.
Opinie na temat odrębności rodzaju różniły się między sobą. John Scannella i Jack Horner uznali go za pośrednie stadium wzrostu pomiędzy triceratopsem i torozaurem[3][84]. Andrew Farke w jego powtórnym opisie jedynej znanej czaszki z 2011 uznał, że chodzi o osobnika w podeszłym wieku reprezentującego jego własny pewny takson, Nedoceratops hatcheri[5]. Nicholas Longrich i Daniel Fields także nie zgodzili się z tezą o stopniowym przejściu między triceratopsem i torozaurem, sugerując, że otwory w kryzie były patologią[7].
Torozaur

Torozaur to rodzaj Ceratopsidae zidentyfikowany na podstawie pary czaszek z 1891, 2 lata po triceratopsie. Przypomina on Triceratops wiekiem geologicznym, występowaniem, anatomią i rozmiarami. Uznano go za bliskiego krewnego triceratopsa[85]. Wyróżnia się z kolei wydłużoną czaszką i obecnością dwóch otworów w kryzie. Paleontolodzy badający ontogenezę (a więc wzrost i rozwój osobnika w czasie jego życia) dinozaurów z formacji Hell Creek w Montanie niedawno zaprezentowali argumenty za uznaniem ich za ten sam rodzaj.
John Scannella w pracy zaprezentowanej w Bristolu w Wielkiej Brytanii na konferencji Society of Vertebrate Paleontology 25 września 2009 reklasyfikował torozaura jako wysoko dojrzałe osobniki triceratopsa, prawdopodobnie należące do tej samej płci. Jack Horner, mentor Scannelli na Montana State University – Bozeman, zwrócił uwagę, że czaszki dinozaurów rogatych budowała kość metaplastyczna. Jej charakterystyczną cechę stanowi wydłużanie się i skracanie z upływem czasu, rozciąga się ona i resorbuje, tworząc nowe kształty. W czaszkach już wcześniej przypisywanych triceratopsowi zauważa się znaczną zmienność. Horner powiedział, że ich rogi odchylają się do tyłu u osobników młodych, stercząc prosto u dorosłych. Szacunkowo połowa wszystkich czaszek dorosłych triceratopsów obejmuje dwa cienkie obszary w kryzie, odpowiadające miejscom, w których u torozaura znajdują się otwory. Ich rozwój zmniejsza masę czaszki, którą z drugiej strony zwiększałoby dojrzewanie triceratopsów połączone z wydłużaniem się kryzy[86]. Publikacja opisująca szczegółowo te wyniki pojawiła się w lipcu 2010. Podpisali się pod nią Scannella i Horner. Argumentowali w niej, że torozaur, podobnie, jak współczesny mu Nedoceratops, stanowią synonimy triceratopsa[3]. Farke (2011) nie zgodził się z ich wnioskami. Zachował odrębność nedoceratopsa. Zaproponował zmiany zachodzące w starzejącym się triceratopsie, które czyniły zeń formę określoną mianem „torozaura”. Wymagały one dodania kości epokcypitalnych, zmiany struktury kości z charakterystycznej dla dorosłych zwierząt na spotykaną u niedojrzałych z następczym powrotem do dojrzałej oraz późny początek pojawiania się dziur w kryzie[5].
Nicholas Longrich i Daniel Field przeprowadzili badanie, w którym nie zgodzili się z wynikami Scanelli i Hornera. Po analizie 35 okazów Longrich i Field doszli do tego, że istniały okazy triceratopsa zbyt stare, by uznać je za młode, oraz torozaury zbyt młode, by uznać je za dorosłe. Stwierdzili również, że jeśliby rzeczywiście triceratops i torozaur były w istocie jednym gatunkiem, to powinny istnieć formy przejściowe pomiędzy nimi. Sugerują jednak, że okaz triceratopsa Scanelli z otworem w kryzie był po prostu chory i nie stanowi formy przejściowej między triceratopsem a torozaurem[7][6].
Ojoceratops i Tatankaceratops
W kolejnej publikacji paleontolog Nick Longrich argumentował, że synonimizacja triceratopsa i torozaura wymaga lepszych form pośrednich, niż wskazane pierwotnie przez Scannellę i Hornera. Longrich zgodził się na uznanie nedoceratopsa za synonim dla Triceratops. Co więcej, zasugerował, że opisany niedawno Ojoceratops był nieodróżnialny od okazu T. horridus przypisanego poprzednio zlikwidowanemu gatunkowi T. serratus. Longrich zauważył, że inny nowo opisany rodzaj, Tatankaceratops, posiada dziwną mieszankę cech młodych i dorosłych okazów triceratopsa. Uznał, że może reprezentować gatunek skarłowaciałego triceratopsa lub po prostu okaz cierpiący na zaburzenia rozwojowe, przez które przedwcześnie zakończył swój wzrost[87].
W kulturze

Wyróżniający się wygląd triceratopsa zapewnił mu częstą obecność w filmie, grach komputerowych i dokumentach. Przykłady stanowią: film z 1993 Park Jurajski i serial paradokumentalny BBC z 1999 Wędrówki z dinozaurami.
Triceratops (bez wyszczególniania gatunku) stanowi skamieniałość stanową Dakoty Południowej[88], jest też oficjalnym dinozaurem stanu Wyoming[89].
Powracającym tematem, szczególnie w książkach dla dzieci, jest pojawiająca się w punkcie kulminacyjnym walka pomiędzy triceratopsem a tyranozaurem. W 1942 Charles R. Knight narysował mural zawierający konfrontację pomiędzy dwoma dinozaurami w Muzeum Historii Naturalnej w Chicago dla National Geographic Society, ustanawiając ich wizerunek w kulturze popularnej jako wrogów[90]. Paleontolog Robert Bakker powiedział o wyobrażonej rywalizacji pomiędzy tyranozaurem i triceratopsem, że żadne starcie pomiędzy drapieżnikiem a jego ofiarą nie było nigdy bardziej dramatyczne[90].
Uwagi
- ↑ W języku polskim zwierzę również określa się mianem triceratopsa, tylko że nazwę polskojęzyczną odmienia się i zapisuje normalną czcionką.
Przypisy
- ↑ O.Ch. Marsh. Notice of gigantic horned Dinosauria from the Cretaceous. „American Journal of Sciences”. Third series. 38, s. 173, 1889. DOI: 10.2475/ajs.s3-38.224.173. (ang.).
- 1 2 G.M. Erickson, K. H. Olson. Bite marks attributable to Tyrannosaurus rex: preliminary description and implications. „Journal of Vertebrate Paleontology”. 16 (1), s. 175–178, 1996. DOI: 10.1080/02724634.1996.10011297.
- 1 2 3 4 5 J. Scannella, J. R. Horner. Torosaurus Marsh, 1891, is Triceratops Marsh, 1889 (Ceratopsidae: Chasmosaurinae): synonymy through ontogeny. „Journal of Vertebrate Paleontology”. 30 (4), s. 1157–1168, 2010. DOI: 10.1080/02724634.2010.483632.
- ↑ Brian Switek: New Study Says Torosaurus=Triceratops. [w:] Dinosaur Tracking [on-line]. Smithsonian.com. [dostęp 2011-03-02].
- 1 2 3 Andrew A. Farke. Anatomy and taxonomic status of the chasmosaurine ceratopsid Nedoceratops hatcheri from the Upper Cretaceous Lance Formation of Wyoming, U.S.A. „PLoS ONE”. 6 (1), s. e16196, 2011. DOI: 10.1371/journal.pone.0016196. PMID: 21283763. PMCID: PMC3024410.
- 1 2 NR Longrich, DJ Field, Torosaurus Is Not Triceratops: Ontogeny in Chasmosaurine Ceratopsids as a Case Study in Dinosaur Taxonomy, „PLoS ONE”, 7 (2), 2012, e32623, DOI: 10.1371/journal.pone.0032623.
- 1 2 3 Neil Bowdler: Triceratops and Torosaurus dinosaurs 'two species, not one’. BBC News, 2012-03-01. [dostęp 2014-02-04]. (ang.).
- ↑ T Dinosaurs Page 2. DinoDictionary.com. [dostęp 2010-08-03]. [zarchiwizowane z tego adresu (2018-10-07)].
- ↑ Triceratops in The Natural History Museum’s Dino Directory. Internt.nhm.ac.uk. [dostęp 2010-08-03]. [zarchiwizowane z tego adresu (2009-02-17)].
- ↑ R.M. Alexander. Mechanics of posture and gait of some large dinosaurs. „Zoological Journal of the Linnean Society”. 83, s. 1–25, 1985. DOI: 10.1111/j.1096-3642.1985.tb00871.x.
- 1 2 D. Lambert: The Ultimate Dinosaur Book. Dorling Kindersley, New York, 1993, s. 152–167. ISBN 1-56458-304-X.
- ↑ S. Perkins, Julius T. Csotonyi. Dressing Up Dinos. „Science News”. 177 (3), s. 22–25, 2010. DOI: 10.1002/scin.5591770321.
- 1 2 3 4 S.-I. Fujiwara. A Reevaluation of the manus structure in Triceratops (Ceratopsia: Ceratopsidae). „Journal of Vertebrate Paleontology”. 29 (4), s. 1136–1147, 2009. DOI: 10.1671/039.029.0406.
- 1 2 3 4 5 Dodson 1996 ↓.
- ↑ P. Christiansen, G.S. Paul. Limb bone scaling, limb proportions, and bone strength in neoceratopsian dinosaurs. „Gaia”. 16, s. 13–29, 2001.
- ↑ S. Thompson, R. Holmes. Forelimb stance and step cycle in Chasmosaurus irvinensis (Dinosauria: Neoceratopsia). „Palaeontologia Electronica”. 10 (1), s. 17, 2007.
- ↑ Habitual locomotor behavior inferred from manual pathology in two Late Cretaceous chasmosaurine ceratopsid dinosaurs, Chasmosaurus irvinensis (CMN 41357) and Chasmosaurus belli (ROM 843). W: E. Rega, R. Holmes, A. Tirabasso: New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium. Michael J. Ryan, Brenda J. Chinnery-Allgeier, David A. Eberth (red.). Bloomington and Indianapolis: Indiana University Press, 2010, s. 340–354. ISBN 978-0-253-35358-0.
- ↑ Martin, A.J.: Introduction to the Study of Dinosaurs. Wyd. 2. Oxford: Blackwell Publishing, 2006. ISBN 1-4051-3413-5.
- ↑ H. G. Liddell, R. Scott: Greek-English Lexicon, Abridged Edition. Oxford University Press, Oxford, UK, 1980. ISBN 0-19-910207-4.
- ↑ triceratops. Online Etymology Dictionary.
- 1 2 3 Hatcher, J. B., Marsh, O. C., & Lull, R. S. (1907) The Ceratopsia. Government Printing Office, Washington, D.C. ISBN 0-405-12713-8.
- ↑ L.M. Lambe. On Eoceratops canadensis, gen. nov., with remarks on other genera of Cretaceous horned dinosaurs. „Canada Department of Mines Geological Survey Museum Bulletin”. 12, s. 1–49, 1915.
- 1 2 R.S. Lull. A revision of the Ceratopsia or horned dinosaurs. „Memoirs of the Peabody Museum of Natural History”. 3 (3), s. 1–175, 1933. [dostęp 2010-11-20].
- 1 2 C.M. Sternberg. The Edmonton fauna and description of a new Triceratops from the Upper Edmonton member; phylogeny of the Ceratopsidae. „National Museum of Canada Bulletin”. 113, s. 33–46, 1949.
- 1 2 Ostrom, J. H.. Functional morphology and evolution of the ceratopsian dinosaurs. „Evolution”. 20 (3), s. 290–308, 1966. DOI: 10.2307/2406631. JSTOR: 2406631.
- ↑ David Norman: The Illustrated Encyclopaedia of Dinosaurs. London: Salamander Books, 1985. ISBN 0-517-46890-5.
- 1 2 3 Lehman, T. M. (1990). The ceratopsian subfamily Chasmosaurinae: sexual dimorphism and systematics. w: Carpenter, K., & Currie, P. J. (red.). Dinosaur Systematics: Perspectives and Approaches. Cambridge University Press, Cambridge, s. 211–229. ISBN 0-521-36672-0.
- ↑ Dodson, P., & Currie, P. J. (1990). Neoceratopsia. 593–618. w: Weishampel, D. B., Dodson, P., & Osmólska, H. (red.). The Dinosauria. University of California Press, Berkeley, s. 593–618. ISBN 0-520-06727-4.
- ↑ P. Dodson. Comparative craniology of the Ceratopsia. „American Journal of Science”. 293, s. 200–234, 1993. DOI: 10.2475/ajs.293.A.200.
- ↑ New Horned Dinosaurs from Utah Provide Evidence for Intracontinental Dinosaur Endimism. „PLoS ONE”, 2010. DOI: 10.1371/journal.pone.0012292. (ang.).
- ↑ J.A. Gauthier. Saurischian monophyly and the origin of birds. The Origin of Birds and the Evolution of Flight, K. Padian (red.). „Memoirs of the California Academy of Sciences”. 8, s. 1–55, 1986.
- ↑ P.C. Sereno. A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria. „Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen”. 210 (1), s. 41–83, 1998.
- ↑ Lehman T.M.. Late Maastrichtian paleoenvironments and dinosaur biogeography in the Western Interior of North America. „Palaeogeography, Palaeoclimatology and Palaeoecology”. 60 (3), s. 290, 1987. DOI: 10.1016/0031-0182(87)90032-0.
- ↑ Dodson 1996 ↓, s. 244.
- 1 2 3 4 5 6 7 8 Dodson, P.; Forster, C.A.; and Sampson, S.D. (2004) Ceratopsidae. In: Weishampel, D. B.; Dodson, P.; & Osmólska, H. (red.), The Dinosauria (2. wydanie). University of California Press, Berkeley, s. 494–513. ISBN 0-520-24209-2.
- 1 2 Joshua C. Mathews, Stephen L. Brusatte, Scott A. Williams, Michael D. Henderson. The first Triceratops bonebed and its implications for gregarious behavior. „Journal of Vertebrate Paleontology”. 29 (1), s. 286–290, 2009. DOI: 10.1080/02724634.2009.10010382.
- ↑ Triceratops trio unearthed in Wyoming. CNN, 2013-06-04. [dostęp 2014-02-04]. (ang.).
- ↑ B.R. Erickson. Mounted skeleton of Triceratops prorsus in the Science Museum. „Scientific Publications of the Science Museum”. 1, s. 1–16, 1966.
- ↑ Dodson 1996 ↓, s. 79.
- ↑ Bakker 1986 ↓, s. 438.
- ↑ Dinosaurs of the Lance Formation in eastern Wyoming. W: K. Derstler, G. E. Nelson (red.): The Dinosaurs of Wyoming. Wyoming Geological Association, 1994, s. 127–146, seria: Wyoming Geological Association Guidebook, 44th Annual Field Conference.
- ↑ J. Tait, B. Brown. How the Ceratopsia carried and used their head. „Transactions of the Royal Society of Canada”. 22, s. 13–23, 1928.
- ↑ J.H. Ostrom. A functional analysis of jaw mechanics in the dinosaur Triceratops. „Postilla”. 88, s. 1–35, 1964. [dostęp 2014-02-04].
- ↑ D.B. Weishampel. Evolution of jaw mechanisms in ornithopod dinosaurs. „Advances in Anatomy, Embryology, and Cell Biology”. 87, s. 1–110, 1984. PMID: 6464809.
- ↑ Coe, M. J.; Dilcher, D. L.; Farlow, J. O.; Jarzen, D. M.; Russell, D. A. (1987). Dinosaurs and land plants. W: Friis, E. M.; Chaloner, W. G.; Crane, P. R. (red.) The Origins of Angiosperms and their Biological Consequences Cambridge University Press, s. 225–258. ISBN 0-521-32357-6.
- ↑ R.S. Lull. The cranial musculature and the origin of the frill in the ceratopsian dinosaurs. „American Journal of Science”. 4 (25), s. 387–399, 1908. DOI: 10.2475/ajs.s4-25.149.387.
- 1 2 Forster, C. A. (1990). The cranial morphology and systematics of Triceratops, with a preliminary analysis of ceratopsian phylogeny. Ph.D. Dissertation. University of Pennsylvania, Philadelphia. s. 227.
- ↑ Bakker 1986 ↓.
- ↑ Sternberg, C. H. (1917). Hunting Dinosaurs in the Badlands of the Red Deer River, Alberta, Canada. Published by the author, San Diego, California, s. 261.
- ↑ An analysis of predator-prey behavior in a head-to-head encounter between Tyrannosaurus rex and Triceratops. W: Happ, J.: Tyrannosaurus rex, the Tyrant King (Life of the Past). Larson, P. & Carpenter, K. (red.). Bloomington: Indiana University Press, 2008, s. 355–368. ISBN 0-253-35087-5.
- ↑ Dodson, Peter, The Horned Dinosaurs, Princeton Press. s. 19.
- ↑ A.A. Farke. Horn Use in Triceratops (Dinosauria: Ceratopsidae): Testing Behavioral Hypotheses Using Scale Models. „Palaeo-electronica”. 7 (1), s. 1–10, 2004. [dostęp 2010-11-20].
- ↑ Tanke, D. H, and Farke, A. A. (2006). Bone resorption, bone lesions, and extracranial fenestrae in ceratopsid dinosaurs: a preliminary assessment. W: Carpenter, K. (red.). Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs Indiana University Press: Bloomington. s. 319–347. ISBN 0-253-34817-X.
- ↑ A.A. Farke, E.D.S. Wolff, D.H. Tanke, Paul Sereno. Evidence of Combat in Triceratops. „PLoS ONE”. 4 (1), s. e4252, 2009. DOI: 10.1371/journal.pone.0004252.
- ↑ JE Peterson, C Dischler, NR Longrich, Distributions of Cranial Pathologies Provide Evidence for Head-Butting in Dome-Headed Dinosaurs (Pachycephalosauridae), „PLoS ONE”, 8 (7), 2013, e68620, DOI: 10.1371/journal.pone.0068620.
- ↑ Michael Wall: Scars Reveal How Triceratops Fought –. Wired.com, 2009-01-27. [dostęp 2010-08-03].
- ↑ Reid REH (1997) Histology of bones and teeth. W: P. J. Currie, K. Padian (red.). Encyclopedia of Dinosaurs. Academic Press, San Diego, CA. 329–339.
- ↑ John R. Horner, Mark B. Goodwin. Extreme Cranial Ontogeny in the Upper Cretaceous Dinosaur Pachycephalosaurus. „PLoS ONE”. 4 (10), s. e7626, 2009-10-27. DOI: 10.1371/journal.pone.0007626. (ang.).
- ↑ Horner JR, Lamm E (2011) Ontogeny of the parietal frill of Triceratops: a preliminary histological analysis. Comptes Rendus Palevol 10: 439–452.
- ↑ Wheeler, P.E.. Elaborate CNS cooling structures in large dinosaurs. „Nature”. 275 (5679), s. 441–443, 1978. DOI: 10.1038/275441a0. PMID: 692723.
- ↑ J. O. Farlow, C. V. Thompson, D. E. Rosner. Plates of the dinosaur Stegosaurus: Forced convection heat loss fins?. „Science”. 192 (4244), s. 1123, 1976. DOI: 10.1126/science.192.4244.1123. PMID: 17748675.
- ↑ Davitashvili, L. Sh.: Teoriya Polovogo Otbora (Theory of Sexual Selection). Izdatel’stvo Akademii nauk SSSR, 1961, s. 538.
- ↑ J.O. Farlow, P. Dodson. The behavioral significance of frill and horn morphology in ceratopsian dinosaurs. „Evolution”. 29 (2), s. 353, 1975. DOI: 10.2307/2407222. JSTOR: 2407222.
- ↑ M.B. Goodwin i inni, The smallest known ''Triceratops'' skull: new observations on ceratopsid cranial anatomy and ontogeny, „Journal of Vertebrate Paleontology”, 26 (1), 2006, s. 103, DOI: 10.1671/0272-4634(2006)26[103:TSKTSN]2.0.CO;2, ISSN 0272-4634 [zarchiwizowane z adresu 2006-09-01].
- ↑ J. O. Farlow, P. Dodson, 1975. The behavioral significance of frill and horn morphology in ceratopsian dinosaurs. Evolution 29: s. 353–361.
- ↑ A. J. Martin (2006). Introduction to the Study of Dinosaurs. Second Edition. Oxford, Blackwell Publishing. s. 299-300. ISBN 1-4051-3413-5.
- 1 2 Carpenter, K. (2006). „Bison” alticornis and O.C. Marsh’s early views on ceratopsians. In: Carpenter, K. (red.). Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs, Indiana University Press, Bloomington and Indianapolis, s. 349–364. ISBN 0-253-34817-X.
- ↑ O.C. Marsh. Notice of new fossil mammals. „American Journal of Science”. 34, s. 323–331, 1887.
- ↑ O.C. Marsh. A new family of horned Dinosauria, from the Cretaceous. „American Journal of Science”. 36, s. 477–478, 1888.
- ↑ O.C. Marsh. Notice of new American Dinosauria. „American Journal of Science”. 37, s. 331–336, 1889a.
- ↑ O.C. Marsh. Notice of gigantic horned Dinosauria from the Cretaceous. „American Journal of Science”. 38, s. 173–175, 1889b.
- ↑ Breithaupt, B.H. (1999). „First Discovery of Dinosaurs in the American West.” s. 59-65 w: Gillette, D.D. (red.), Vertebrate Paleontology In Utah. Utah Geological Survey. ISBN 1-55791-634-9, ISBN 978-1-55791-634-1.
- ↑ Morph-osaurs: How shape-shifting dinosaurs deceived us – life – 28 July 2010. New Scientist. [dostęp 2010-08-03].
- ↑ J.H. Ostrom, P. Wellnhofer. The Munich specimen of Triceratops with a revision of the genus. „Zitteliana”. 14, s. 111–158, 1986.
- ↑ Loxodonta cyclotis, [w:] Integrated Taxonomic Information System [dostęp 2022-12-30] (ang.).
- ↑ C.A. Forster. Species resolution in Triceratops: cladistic and morphometric approaches. „Journal of Vertebrate Paleontology”. 16 (2), s. 259–270, 1996. DOI: 10.1080/02724634.1996.10011313.
- ↑ T.M. Lehman. A gigantic skull and skeleton of the horned dinosaur Pentaceratops sternbergi from New Mexico. „Journal of Paleontology”. 72 (5), s. 894–906, 1998. JSTOR: 1306666.
- ↑ J. B. Scannella, D. W. Fowler (2009). „Anagenesis in Triceratops: evidence from a newly resolved stratigraphic framework for the Hell Creek Formation.” s. 148–149 w 9th North American Paleontological Convention Abstracts. Cincinnati Museum Center Scientific Contributions 3.
- ↑ John Bell Hatcher. Two new Ceratopsia from the Laramie of Converse County, Wyoming. „American Journal of Science”. 4 (Series 4), s. 413−419, 1905.
- ↑ Richard Swann Lull. Restoration of the horned dinosaur Diceratops. „American Journal of Science”. 4 (Series 4), s. 420−422, 1905.
- ↑ A. S. Ukrainsky (2007). „A new replacement name for Diceratops Lull, 1905 (Reptilia: Ornithischia: Ceratopsidae).” Zoosystematica Rossica, 16(2), 20 grudnia 2007: 292.
- ↑ Octávio Mateus. Two ornithischian dinosaurs renamed: Microceratops Bohlin 1953 and Diceratops Lull 1905. „Journal of Paleontology”. 82 (2), s. 423, 2008. DOI: 10.1666/07-069.1.
- ↑ A. S. Ukrainsky (2009). „Sinonimiya rodov Nedoceratops Ukrainsky, 2007 i Diceratus Mateus, 2008 (Reptilia: Ornithischia: Ceratopidae).” Paleontologicheskii zhurnal, 2009(1): 108. Tłumaczenie na jęz. ang.: A. S. Ukrainsky (2009). „Synonymy of the genera Nedoceratops Ukrainsky, 2007 and Diceratus Mateus, 2008 (Reptilia: Ornithischia: Ceratopidae).” Paleontological Journal, 2009 43(1): s. 116.
- ↑ J.B. Scannella, J.R. Horner. ‘Nedoceratops’: An Example of a Transitional Morphology. „PLoS ONE”. 6 (12), s. e28705, 2011. DOI: 10.1371/journal.pone.0028705. (ang.).
- ↑ A. A. Farke: Cranial osteology and phylogenetic relationships of the chasmosaurine ceratopsid Torosaurus latus. W: K. Carpenter: Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs. Bloomington: Indiana University Press, 2006, s. 235–257. ISBN 978-0-253-34817-3.
- ↑ New Analyses Of Dinosaur Growth May Wipe Out One-third Of Species. [w:] Science News [on-line]. ScienceDaily.com, 2009-10-31. [dostęp 2009-11-03].
- ↑ Nicholas R. Longrich. Titanoceratops ouranous, a giant horned dinosaur from the Late Campanian of New Mexico. „Cretaceous Research”. 32 (3), s. 264–276, 2011. DOI: 10.1016/j.cretres.2010.12.007.
- ↑ South Dakota Department of Tourism: South Dakota Facts. [dostęp 2014-02-04].
- ↑ State of Wyoming: State of Wyoming – General Information. [dostęp 2007-01-20]. [zarchiwizowane z tego adresu (2007-02-10)].
- 1 2 Bakker, R.T. 1986. The Dinosaur Heresies. New York: Kensington Publishing, s. 240.
Bibliografia
- R.T. Bakker, The Dinosaur Heresies: New Theories Unlocking The Mystery of the Dinosaurs and Their Extinction, New York: William Morrow, 1986, ISBN 0-14-010055-5.
- P. Dodson, The Horned Dinosaurs, Princeton, New Jersey: Princeton University Press, 1996, ISBN 0-691-02882-6.
- kategoria
- klasyfikacja
- lista
- wikiprojekt
| Infrarzędy dinozaurów | |
|---|---|
| Era dinozaurów | |
| Dyscypliny powiązane |
