| Paraceratherium | |||||
| Forster-Cooper, 1911 | |||||
| Okres istnienia: 34–23 mln lat temu | |||||
 Odlew szkieletu P. transouralicum, Narodowe Muzeum Przyrody i Nauki (Tokio) | |||||
| Systematyka | |||||
| Domena | |||||
|---|---|---|---|---|---|
| Królestwo | |||||
| Typ | |||||
| Podtyp | |||||
| Gromada | |||||
| Podgromada | |||||
| Infragromada | |||||
| Rząd | |||||
| Rodzina | |||||
| Podrodzina | |||||
| Rodzaj |
Paraceratherium | ||||
| |||||
| Gatunki | |||||
| |||||
| Zasięg występowania | |||||
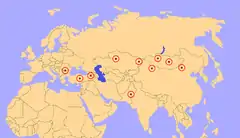 Miejsca, gdzie znajdywano skamieniałości Paraceratherium | |||||
Paraceratherium – wymarły rodzaj bezrogiego nosorożca, a właściwie przedstawiciel rodziny Hyracodontidae z podrodziny Indricotheriinae; jeden z największych ssaków lądowych w historii. Żył od wczesnej do późnej epoki oligoceńskiej (34–23 miliony lat temu). Jego szczątki znaleziono w Eurazji od Chin do terenów byłej Jugosławii. Nazwa Paraceratherium oznacza „obok bezrogiej bestii”, odnosi się do Aceratherium, rodzaju uznawanego niegdyś za podobny.
Dokładna wielkość Paraceratherium pozostaje nieznana z powodu niekompletności znalezisk. Masę ciała zwierzęcia szacuje się na 15-20 ton. Wysokość w kłębie wynosiła około 4,8 m, długość zaś około 7,4 m. Długie kończyny przypominały słupy. Długa szyja nosiła głowę o czaszce mierzącej 1,3 m. Długie siekacze przypominały ciosy. Wcięcie nosowe sugeruje obecność chwytnej górnej wargi bądź nawet trąby.
Trybem życia Paraceratherium mogło przypominać współczesne wielkie ssaki, jak słoniowate czy przetrwałe do dziś nosorożcowate. Z uwagi na rozmiary groziło mu niewielu drapieżników, natomiast rozmnażanie następowało wolno. Paraceratherium żywiło się liśćmi, miękkimi roślinami, krzewami. Zwierzęta te zasiedlały zróżnicowane siedliska – od suchych pustyń porosłych nielicznymi drzewami, aż do podzwrotnikowych lasów. Przyczyny wyginięcia tych zwierząt nie są znane, zaproponowano różne czynniki mogące odgrywać tu rolę.
Taksonomia rodzaju i zaliczanych doń gatunków ma bogatą i skomplikowaną historię. Opisywano inne rodzaje Indricotheriinae, jak Baluchitherium, Indricotherium i Dzungariotherium. Nie znaleziono jednak kompletnych okazów, co utrudnia porównania i klasyfikację. Większość współczesnych naukowców uznaje wymienione rodzaje za młodsze synonimy Paraceratherium, w obrębie którego wyróżnia się 4 gatunki: P. bugtiense (gatunek typowy), P. transouralicum, P. prohorovi i P. orgosensis, choć ten ostatni może stanowić odrębny rodzaj. Najlepiej z nich poznano P. transouralicum, które przedstawia większość rekonstrukcji rodzaju. Różnice pomiędzy P. bugtiense i P. transouralicum mogą być spowodowane w rzeczywistości dymorfizmem płciowym, co czyniłoby z tych dwojga jeden gatunek.
Historia badań i taksonomia
Historia taksonomiczna Paraceratherium jest złożona z powodu fragmentaryczności znalezionych szczątków, a także izolacji zachodnich, radzieckich i chińskich naukowców – przez większość XX wieku pracujących osobno i publikujących swe odkrycia głównie w swych rodzimych językach[1]. Uczeni z różnych części świata starali się porównać swe wyniki, by znaleźć bardziej kompletny obraz tych stworzeń, jednakże utrudniały im to polityka i wojny[2]. Przeciwstawne tendencje taksonomiczne (ang. lumping and splitting, klasyfikowanie z użyciem mniejszych bądź większych kategorii) również przyczyniły się do problematycznej sytuacji[3]. Niedokładne datowanie geologiczne doprowadziło wcześniej naukowców do poglądu, jakoby różne formacje geologiczne, obecnie uznawane za równowiekowe, powstawały w różnym czasie. Wiele nazw rodzajowych utworzono na podstawie subtelnych różnic trzonowców, różniących się pomiędzy populacjami innych taksonów Rhinocerotoidea, większość uczonych nie akceptuje ich jako cech stanowiących o wyróżnianiu gatunków[4].
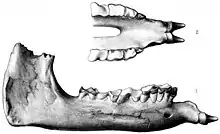
Wczesne odkrycia Indricotheriinae nauka zawdzięcza kolonialnym związkom z Azją[2]. Pierwsze znane szczątki Indricotheriinae zebrał w Beludżystanie (współczesny Pakistan) w 1846 żołnierz Vickary, wtedy jednak nie potrafiono ich identyfikować[5]. Pierwsze skamieliny obecnie zaliczane do Paraceratherium odkrył brytyjski geolog Guy Ellcock Pilgrim w Balochistanie w latach 1907–1908. Zebrany przezeń materiał obejmował szczękę, zęby żuchwy i jej tylny fragment. Pochodził on z formacji Chitarwata z Dera Bugti, gdzie Pilgrim prowadził swe badania. W 1908 na podstawie tych pozostałości opisał nowy gatunek nosorożca z rodzaju Aceratherium: A. bugtiense. Aceratherium służył jako takson-worek na śmieci, obejmował kilka niespokrewnionych bliżej gatunków bezrogich nosorożców, z których wiele przeniesiono potem do innych rodzajów[1][6]. Skamieniałe siekacze przypisane przez Pilgrima do niespokrewnionego z wyżej wymienionym rodzaju Bugtitherium okazały się później należeć do tego nowego gatunku[7].
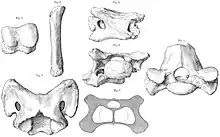
W 1910 odkryto więcej niekompletnych skamielin w Dera Bugti, podczas wyprawy brytyjskiego paleontologa Clive’a Forstera-Coopera. Na podstawie tych szczątków Foster-Cooper przeniósł A. bugtiense do nowego rodzaju Paraceratherium, którego nazwa znaczy „obok bezrogiej bestii”, odnosząc się do Aceratherium[1][8]. Reklasyfikację uzasadniał wydatnie zakręcającymi w dół dolnymi ciosami zwierzęcia[7]. W 1913 Forster-Cooper nazwał nowy rodzaj i gatunek Thaumastotherium osborni. Nazwa rodzajowa oznacza „wspaniałą bestię”. Uczony bazował na większych skamieniałościach z tych samych wykopalisk, jednak zmienił nazwę na Baluchitherium później tego samego roku, gdyż poprzedniej użyto już dla określenia owada z grupy pluskwiaków[9][10]. Skamieniałości Baluchitherium były tak fragmentaryczne, że Foster-Cooper był zdolny stwierdzić jedynie, że zwierzę należy do nieparzystokopytnych, wspomniał jednak możliwość pomyłki z Paraceratherium[11]. Amerykański paleontolog Henry Fairfield Osborn, na cześć którego nazwano B. osborni, zasugerował, że może należeć do Titanotheriidae[2].
Ekspedycja Rosyjskiej Akademii Nauk znalazła później skamieliny w formacji Aral w pobliżu Jeziora Aralskiego w Kazachstanie. Był to najbardziej kompletny znany szkielet Indricotheriinae, brakowało mu jednak czaszki. W 1916, bazując na tych szczątkach, Aleksiej Aleksiejewicz Borissiak wskrzesił rodzaj Indricotherium, nazwany po mitologicznym potworze, Indriku. Nie przypisał mu epitetu gatunkowego aż do 1923, gdy utworzył nazwę I. asiaticum, jednak Maria Pawłowa zdążyła już nazwać je I. transouralicum w 1922[1][12]. Również w 1923 Borissiak utworzył nową podrodzinę Indricotheriinae, by objąć nią różne spokrewnione ze sobą znane wtedy formy[13]. W 1939 Borissiak również nazwał nowy gatunek Paraceratherium z Kazachstanu, P. prohorovi[14].
W 1922 amerykański odkrywca Roy Chapman Andrews poprowadził dobrze udokumentowaną wyprawę do Chin i Mongolii, sponsorowaną przez Amerykańskie Muzeum Historii Naturalnej. Różne szczątki Indricotheriinae odnaleziono w formacjach mongolskiej pustyni Gobi, w tym kończyny osobnika stojącego w pozycji wyprostowanej, co wskazuje, że umarł on w kurzawce. Zalicza się doń również czaszka o dużym stopniu kompletności. Pozostałości te stały się podstawą opisu Baluchitherium grangeri, nazwanego przez Osborna w 1923[15][16].
Dzungariotherium orgosensis opisano w 1973 głównie w oparciu o zęby z Dżungarii w Sinciang na północnym zachodzie Chin[17]. Na podstawie kolejnych szczątków, kierując się różnicami w wielkości, kształcie pyska i ułożeniu przednich zębów ukuto wiele innych nazw rodzajowych i gatunkowych. Skamieniałości przypisywane Paraceratherium cały czas odkrywa się w Eurazji, jednak sytuacja polityczna w Pakistanie stała się zbyt niestabilna, by prowadzić w tym kraju dalsze wykopaliska[5].
Gatunki i synonimy
W 1936 amerykańscy paleontolodzy Walter Granger i William K. Gregory zaproponowali uznanie Forster-Cooperowego Baluchitherium osborni za prawdopodobnie młodszy synonim Paraceratherium bugtiense, ponieważ okazy zebrano w tej samej lokalizacji i należały prawdopodobnie do tego samego gatunku wykazującego zmienność morfologiczną[18]. William Diller Matthew i Forster-Cooper wyrażali podobne wątpliwości kilka lat wcześniej. Choć uznano ją już za młodszy synonim, nazwa rodzajowa Baluchitherium stała się popularna w różnych mediach dzięki rozgłosowi wokół osbornowego B. grangeri[3][19].

W 1989 paleontolodzy Spencer G. Lucas i Jay C. Sobus opublikowali rewizję taksonów Indricotheriinae, którą popiera do dziś większość zachodnich specjalistów. Autorzy przedstawili konkluzję, że Paraceratherium, jako najstarsza z nazw, stanowi jedyny pewny rodzaj Indricotherinae z oligocenu, zawierając 4 pewne gatunki: P. bugtiense, P. transouralicum, P. prohorovi i P. orgosensis. Wiele innych nazw uznano za młodsze synonimy tych taksonów bądź też nomina dubia bazujące na szczątkach zbyt fragmentarycznych, by umożliwiały dokładną identyfikację. Analiza rzekomych różnic pomiędzy nazwanymi rodzajami i gatunkami Lucasa i Sobusa pokazała, że najprawdopodobniej reprezentują one zmienność pomiędzy populacjami, a większość cech była nie do odróżnienia pomiędzy osobnikami, jak zauważono w latach trzydziestych XX wieku. Fakt, że jedna czaszka, przypisywana do P. transouralicum lub Indricotherium, była kopulasta, podczas gdy inne miały płaskie sklepienie, przypisano dymorfizmowi płciowemu. Możliwe jest więc, że skamieniałości opisywane jako P. bugtiense reprezentują w rzeczywistości samice, podczas gdy P. transouralicum są samcami tego samego gatunku[3][14].
Wedle Lucasa i Sobusa gatunek typowy P. bugtiense z późnego oligocenu z Pakistanu ma następujące młodsze synonimy: B. osborni i P. zhajremensis. P. transouralicum, wcześniej Indricotherium, z późnego oligocenu z terenów Kazachstanu, Mongolii i północnych Chin obejmuje B. grangeri i I. minus. P. orgosensis, wcześniej Dzungariotherium, ze środkowego i późnego oligocenu z północnego zachodu Chin ma D. turfanensis i P. lipidus[14]. P. orgosensis może wykazywać wystarczającą odrębność dla własnej nazwy rodzajowej, jednak jego właściwa pozycja wymaga jeszcze oceny. P. prohorovi z późnooligoceńskiego Kazachstanu może być reprezentowany przez zbyt mało kompletny materiał dla ustalenia jego pozycji. To samo odnosi się do zaproponowanych gatunków I. intermedium i P. tienshanensis, jak również dla rodzajów Benaratherium i Caucasotherium[3][14]. Choć nazwa rodzajowa Indricotherium stanowi obecnie młodszy synonim Paraceratherium, podrodzina zachowała swą nazwę Indricotheriinae, ponieważ synonimizacja nazwy rodzajowej nie pociąga za sobą zmiany nazw wyższych taksonów, nawet jeśli pochodzą one od niej[20].
W przeciwieństwie do rewizji Lucasa i Sobus, publikacja chińskich badaczy z 2003 sugeruje uznanie Indricotherium i Dzungariotherium za pewne rodzaje, w związku z czym P. prohorovi nie należałby do Paraceratherium. Uznają także takie gatunki, jak P. lipidus, P. tienshanensis i P. sui[21]. Praca z 2004 napisana przez chińskiego paleontologa nazwiskiem Tao Deng i współpracowników podaje trzy odrębne rodzaje[22]. Niektórzy autorzy zachodni także używają nazw uznawanych za niepewne od rewizji 1989, nie podając jednak drobiazgowej analizy i nie próbując uzasadniać takiego stanowiska[3].
Ewolucja


Nadrodzina Rhinocerotoidea, do której zaliczają się też współczesne nosorożce, sięga wstecz przynajmniej do wczesnego eocenu, 50 milionów lat temu, kiedy to żył Hyrachyus. Rhinocerotoidea obejmują 3 rodziny: Amynodontidae, Rhinocerotidae (nosorożcowate) i Hyracodontidae. Różnorodność w tej grupie była w czasach prehistorycznych dużo większa. Rozmiary wahały się od rozmiarów psa aż do Paraceratherium. Były to długonogie formy dobrze przystosowane do biegu i przysadziste zwierzęta prowadzące częściowo wodny tryb życia. Większość gatunków nie miało rogów. Ich skamieliny identyfikuje się głównie dzięki cechom zębów, które zachowują się najlepiej z całego szkieletu. Górne trzonowce większości z nich noszą π-kształtny wzór na koronie. Na każdym dolnym zębie trzonowym widnieje z kolei podwójny kształt L. Różnorodne cechy czaszek także służą ich identyfikacji[23].
Podrodzina Indricotheriinae, do której zalicza się Paraceratherium, została po raz pierwszy uwzględniona jako część rodziny Hyracodontidae przez Leonarda B. Radinsky’ego w 1966. Wcześniej traktowano ją jako podrodzinę wśród Rhinocerotidea, a nawet osobną rodzinę Indricotheriidae[24]. W wydanym w 1999 badaniu kladystycznym Tapiromorpha Luke Holbrook umieścił indrikoteria poza kladem Hyracodontidae, pisząc też, że nie muszą one być grupą monofiletyczną[25]. Ustalenia Radinsky’ego stanowią obecnie przeważającą hipotezę. Rodzina Hyracodontidae obejmuje długonogie ssaki zaadaptowane do biegu, takie jak Hyracodon. Odróżniają się od innych budową siekaczy. Indricotheriinae wyróżniają się wśród nich większymi rozmiarami oraz synapomorfiami pyska, siekaczy i kłów. Najwcześniejszym znanym przedstawicielem indrikoteriów jest osiągająca rozmiary psa, żyjąca w środkowym i późnym eocenie w zachodniej Ameryce Północnej i w Azji. Juxia wielkości krowy żyła w środkowym eocenie. W późnym eocenie azjatyckie Urtinotherium osiągnęło rozmiary zbliżające się do Paraceratherium[23][14]. Paraceratherium zamieszkiwało Eurazję w epoce oligocenu, pomiędzy 23 i 34 miliony lat temu[26]. Rodzaj wyróżnia się wśród Indricotheriinae wielkim wzrostem, wcięciem nosowym wspierającym umięśniony pysk i zawijającą się ku dołowi kością przysieczną[14]. Straciło także 2. i 3. siekacz dolny, dolny kieł, jak też 1 dolny przedtrzonowiec[23].
Poniższy kladogram pochodzi z rewizji indrikoteriów z 1989 autorstwa Lucasa i Sobusa[14]:
| Hyracodontidae |
| ||||||||||||||||||||||||
Lucas i współpracownicy doszli do podobnych wniosków w swej poprzedniej analizie Forstercooperia z 1981. Pozostawili weń Paraceratherium i Indricotherium jako osobne rodzaje[27].
Budowa
Wielkość

Paraceratherium uważa się za największego lądowego ssaka, jaki kiedykolwiek istniał. Jednak jego dokładna wielkość nie jest znana z uwagi na niekompletność materiału kopalnego. Wczesne oszacowania masy na 30 ton uważa się obecnie za przesadzone. Zwierzę mogło ważyć maksymalnie 15-20 ton, średnio zaś tylko 11 ton. Obliczenia bazowały głównie na skamielinach P. transouralicum, jako że gatunek ten cechuje się najbardziej kompletnymi szczątkami[4]. Szacunki opierały się o pomiary czaszki, zęby i kości kończyn, jednak znane pozostałości kości pochodzą od osobników różnej wielkości, wobec czego wszystkie rekonstrukcje szkieletu stanowią złożone ekstrapolacje. Skutkuje to różnymi zakresami oszacowanej masy[28][29]. Całkowitą długość ciała Granger i Gregory w 1936 oszacowali na 8,7 m, natomiast Gromova w 1959 na 7,4 m. Wcześniejsze przybliżenie uznaje się dziś za przeszacowane. Do masy ciała Paraceratherium zbliżały się, a być może przewyższały ją, pewne wymarłe trąbowce, o czym świadczy m.in. największy kompletny szkielet mamuta stepowego (Mammuthus trogontherii)[28] czy niekompletne szczątki mastodontów Mammut borsoni i słoni Palaeoloxodon namadicus[30]. Pomimo mniej więcej podobnych mas Paraceratherium było jednak wyższe, niż jakikolwiek trąbowiec[4]. Wysokość w kłębie szacują na 5,25 m Granger i Gregory, na 4,8 m zaś Gregory S. Paul w 1997[31]. Długość szyi szacuje się na 2–2,5 m, jak piszą Michael P. Taylor i Mathew J. Wedel w 2013[32]. Zęby P. orgosensis (to głównie z nich znany jest ten gatunek) są o 25% większe, niż należące do P. transouralicum, co czyni go największym ze znanych indrikoteriów[3].
Tułów

Nie znaleziono jeszcze kompletnego zbioru kręgów ani żeber Paraceratherium. Kompletnie nieznany pozostaje ogon. Dźwigacz i obrotnik są szersze, niż u większości współczesnych nosorożców. Pozostawia to przestrzeń na silne więzadła i mięśnie, potrzebne do utrzymania dużej głowy. Pozostałe kręgi także były bardzo szerokie, miały szerokie wyrostki stawowe, pozostawiające dużo miejsca dla mięśni, ścięgien, więzadeł i nerwów, by wspierać głowę, szyję i kręgosłup. Długie wyrostki kolczyste tworzyły długi garb na grzbiecie, gdzie przyczepiały się mięśnie szyi i więzadło karkowe utrzymujące czaszkę. Żebra przypominały spotykane u współczesnych nosorożców, jednak klatka piersiowa pośród długich kończyn i wielkiego ciała wydawała się względnie mniejsza. Dzisiejsze nosorożcowate cechują się bowiem względnie krótszymi kończynami. Ostatni kręg lędźwiowy łączył się z kością krzyżową. Jest to cecha spotykana u zaawansowanych Rhinocerotoidea[4]. Podobnie jak u zauropodów w kręgach przedkrzyżowych występowały otwory, przypominające pleurocele. Brały one udział w redukcji masy szkieletu[33].
Kończyny były duże i silnie zbudowane, wspierając znaczną masę zwierzęcia. Przypominały pod pewnymi względami i wykazywały konwergencję z kończynami słoni czy zauropodów – zwierząt ciężkich i nieporuszających się zbyt szybko. W przeciwieństwie do tych zwierząt, wykazujących tendencję do wydłużania kości górnych kończyn i skracania, łączenia i kompresji dolnych, dłoni i stóp, Paraceratherium cechowało się krótkimi kośćmi górnymi kończyn, długimi zaś kośćmi dłoni i stóp, nie licząc dyskoidalnych paliczków. Podobny układ występował u biegających Rhinocerotoidea, od których indrikoteria pochodzą. Pewne kości stóp mierzyły prawie 50 cm długości. Kość udowa typowo mierzyła 1,5 m długości. Większe kości udowe spotykano jedynie u niektórych słoni i dinozaurów. Kość udowa jest słupowata, dużo grubsza i znacznie potężniejsza, niż u innych Rhinocerotoidea. Trzy krętarze po bokach uległy znacznej redukcji, jako że mocna budowa kości zmniejsza ich znaczenie. Kończyny zwierzę trzymało prosto jak kolumny, a nie w zgięciu, jak mniejsze zwierzęta, co zmniejszało wymagania stawiane mięśniom kończyn[4]. Przednie łapy kończyły się trzema palcami[34]

Z powodu fragmentarycznej natury znanych skamieniałości Paraceratherium zwierzę od czasu jego odkrycia rekonstruowano na kilka odmiennych sposobów[35]. W 1923 W. D. Matthew nadzorował artystę rysującego taką rekonstrukcję szkieletu w oparciu o okazy jeszcze mniej kompletne, niż już wtedy znane. Wykorzystano proporcje dzisiejszych nosorożcowatych[36]. Rezultat był zbyt przysadzisty, zbity. Osborn później w tym samym roku dysponował już bardziej smukłą rekonstrukcją. Pewne późniejsze przedstawienia czyniły z kolei zwierzę zbyt szczupłe, zwracając niewielką uwagę na szkielet[4]. Gromova opublikowała bardziej kompletną rekonstrukcję szkieletu w 1959, bazując na kośćcu P. transouralicum z formacji Aral. Także brakowało mu kilku kręgów szyjnych[37].
Nic nie wiadomo o kolorze i teksturze skóry zwierzęcia. Nie odkryto bowiem żadnych jej odcisków ani mumii. Większość zażyciowych rekonstrukcji przedstawia zwierzę w grubej, pofałdowanej, szarej, bezwłosej skórze, bazując na współczesnych nosorożcach. Ponieważ włosy zatrzymują ciepło produkowane przez organizm, współczesne wielkie ssaki, jak słonie czy nosorożcowate, są w dużym stopniu pozbawione włosia. Amerykański paleontolog Donald Prothero zaproponował, w przeciwieństwie do większości przedstawień, wygląd Paraceratherium z dużymi, przypominającymi słoniowe uszami służącymi termoregulacji. Małżowiny uszne słoniowatych powiększają powierzchnię ciała. Są dobrze ukrwione, co dodatkowo ułatwia rozpraszanie ciepła. Zgodnie z poglądem Prothero podobnie wyglądało to u Paraceratherium; zwrócił on uwagę na dobrze zbudowane kości wokół otworów słuchowych zewnętrznych[4]. Paleontolog Pierre-Olivier Antoine i Darren Naish odnieśli się do tego pomysłu sceptycznie[38][39].
Czaszka
Największe czaszki Paraceratherium mierzą około 1,3 m długości, 33–38 cm w tylnej części czaszki, a 61 cm szerokości w przekroju poprzez łuki jarzmowe. Paraceratherium miało wysokie czoło, smukłe i pozbawione chropowatego obszaru służącego za punkt przyczepu rogów innych nosorożców. Kości ponad okolicą nosową są długie. Wcięcie nosowo-siekaczowe sięga daleko w obręb czaszki. Wskazuje to, że Paraceratherium dysponowało chwytną górną wargą, podobną do spotykanej u nosorożca czarnego czy indyjskiego, lub nawet krótką trąbą, jak tapirowate. Tylna część czaszki była niska i wąska, pozbawiona dużego grzebienia węgłowego na szczycie czaszki i grzebienia strzałkowego, obserwowanych u rogatych i zaopatrzonych w ciosy zwierząt, potrzebujących silnych mięśni, by pchać i walczyć. Głębokie zagłębienie służyło za to przyczepowi więzadła karkowego, automatycznie podnoszącego czaszkę. Kłykieć potyliczny jest szeroki, Paraceratherium wydaje się posiadać duże, silne mięśnie szyi, pozwalające zwierzęciu obracać głowę silnie w dół przy żerowaniu na gałązkach[4]. Jedna z czaszek P. transouralicum cechuje się kopulastym czołem, podczas gdy u innych jest ono płaskie. Prawdopodobnie odpowiada za to dymorfizm płciowy[14]. Odlew mózgu P. transouralicum wskazuje, że narząd ten obejmował jedynie 8% długości czaszki. U nosorożca indyjskiego wartość ta wynosi 17,7%[18].
Gatunki Paraceratherium rozróżniane są na podstawie cech czaszki. P. bugtiense i P. orgosensis dzielą takie cechy, jak względnie smukłe kość szczękowa i kość przedszczękowa, płytkie sklepienie czaszki, wyrostki sutkowato-przypotyliczne względnie cienkie, położone z tyłu czaszki, grzebień węgłowy sięgający w mniejszym stopniu ku tyłowi, kłykieć potyliczny ułożony poziomo. P. transouralicum miało dobrze zbudowane kości szczękową i przedszczękową, kości jarzmowe kierujące się ku górze, kopulaste kości czołowe, gruby wyrostek sutkowato-przypotyliczny, grzebień węgłowy rozpościerający się do tyłu i kłykcie potyliczne o ułożeniu pionowym[3]. P. orgosensis wyróżnia się od innych gatunków większymi rozmiarami zębów i pewnymi cechami trzonowców[14].
W przeciwieństwie do najprymitywniejszych Rhinocerotoidea przednie zęby Paraceratherium uległy redukcji do pojedynczej pary siekaczy w żuchwie i szczęce. Były one duże i stożkowate, opisuje się je jako ciosy. Górne siekacze kierowały się do dołu. Dolne były krótsze, kierowały się ku przodowi[4]. Cechowały się stożkowatą koroną i prostym korzeniem, korzenie zębów w przekroju różniły się u różnych osobników[34]. Taki układ jest unikalny wśród Rhinocerotoidea dla Paraceratherium i spokrewnionego zeń Urtinotherium. Siekacze mogły być większe u samców. Kły zostały utracone. Siekacze były oddzielone od zębów policzkowych dużą diastemą[4]. Cechę tę spotyka się u ssaków, u których siekacze i zęby policzkowe wyspecjalizowały się w różnych kierunkach[23]. Przedtrzonowce tylko częściowo zachowały wzór π. Każdy z trzonowców osiągał wielkość ludzkiej pięści. Przekraczają je rozmiarami jedynie zęby trąbowców[4]. Pokrywało je grube szkliwo[34]. Górne trzonowce, z wyjątkiem V-kształtnego górnego trzonowca, cechują się π-kształtnym wzorem i zredukowanym metastylem. Zęby policzkowe były L-kształtne, co jest typowe dla Rhinocrotoiea[4]. Trzonowce miały po 3 korzenie: przedni, tylny i wargowy[34].
Paleobiologia

Zoolog Robert M. Alexander zasugerował, że przegrzewanie się mogło stanowić dla Paraceratherium poważny problem z uwagi na jego wielkość[40]. Zgodnie z Prothero analogii należy szukać wśród dużych ssaków, jak słonie, nosorożcowate i hipopotamowate. By wspomóc termoregulację, zwierzęta te za dnia spoczywają w cieniu lub tarzają się w błocie lub wodzie. Żerują i przemieszczają się głównie w nocy. Z powodu swych dużych rozmiarów Paraceratherium nie potrafiło biegać ani w ogóle poruszać się szybko. Potrafiło jednak przemierzać duże dystanse, co było konieczne w środowisku o skąpych zasobach żywieniowych. Mogły występować szerokie areały osobnicze, a także migracje[4]. Prothero sugerował, że zwierzęta tak duże jak indrikoteria potrzebowały bardzo rozległych areałów osobniczych bądź terytoriów, przynajmniej 1 tys. km², i z powodu ubogich zasobów żywnościowych było w Azji mało miejsc dla dużej liczby populacji bądź licznych prawie nieróżniących się od siebie gatunków i rodzajów. Zachodzi tu bowiem zasada Gausego, tłumacząca także to, że nosorożec czarny odżywiający się wyżej rosnącą roślinnością niż trawożerny nosorożec biały może zamieszkiwać te same tereny w Afryce – gatunki te zajmują różne nisze ekologiczne[3].
Większość drapieżników w ich siedliskach osiągało względnie małe rozmiary – mniej więcej wielkości wilka – nie zagrażało więc Paraceratherium[26]. Dorosłe osobniki były zbyt duże dla większości drapieżników, by mogły je zaatakować. Jednak młode były podatne na ataki drapieżników. Ślady ugryzień na kościach z pokładów z Bugti wskazują jednak, że nawet dorosłe zwierzęta stawały się ofiarami ataków mierzących 10–11 m krokodyli, Crocodylus bugtiensis. Jak w przypadku słoni, ciąża mogła u Paraceratherium trwać długo. Zwierzęta te były też długowieczne[4]. Paraceratherium mogły żyć w niewielkich stadach, składających się z samic i ich cieląt, co chroniłoby je przed drapieżnikami[26]. Jako maksymalną masę osiągalną przez ssaki lądowe zaproponowano 20 ton i zbliżało się do tej granicy[41]. Powody, dla których ssaki nie mogą osiągać większych rozmiarów jak zauropody, pozostają nieznane. Powody te mogą wynikać nie z biomechaniki, ale z ekologii, być może wiążą się ze strategią rozrodu[28]. Ruchy, dźwięki i inne zachowania przedstawiane w dokumentach typu CGI, jak Wędrówki z bestiami, są całkowicie hipotetyczne[4].
Pożywienie

Proste zęby o niskich koronach wskazują, że Paraceratherium żywiło się wyżej rosnącą roślinnością. Spożywało miękkie liście i krzewy. Późniejsze Rhinocerotoidea żywiły się niską, trawiastą roślinnością. Posiadały zęby o wysokich koronach, gdyż ich dieta zawierała drobiny gleby, na których ścierały się zęby żuchwy. Badania średniej wielkości fasetek zębów (mesowear) potwierdziły, że Paraceratherium żywiły się miękkim pokarmem – liśćmi. Badania mikrofasetek dopiero czekają na przeprowadzenie[4]. Analiza izotopowa wykazała, że Paraceratherium pasł się głównie na roślinach przeprowadzających fotosyntezę C3, żywiąc się głównie liśćmi[42][43]. Jak jego krewni spośród nieparzystokopytnych – konie, tapiry, nosorożce – Paraceratherium przeprowadzało fermentację w dystalnym jelicie. Pozwalało to na wykorzystanie względnie małych ilości substancji odżywczych z pożywienia, musiało więc pobierać jego znaczne ilości. Paraceratherium powinno więc mieć długi przewód pokarmowy[4].
Granger i Gregory argumentowali, że duże siekacze służyły do obrony bądź do odzierania krzaków poprzez schylenie głową, jakby przy użyciu kilofów i dźwigni[18]. Tapirowate wykorzystują swe trąby, owijając je wokół gałęzi, gdy odkorowują je przednimi zębami. Umiejętność taka przydawałaby się Paraceratherium. Niektórzy autorzy rosyjscy sugerowali, jakoby ciosy służyły do łamania gałęzi, odkorowywania i zginania ich i dlatego też gatunek ze wczesnego oligocenu miał większe ciosy, niż późniejsze, opierając swą dietę w większym stopniu na korze niż na liściach. Jednakże obecnie oba gatunki uważa się za współczesne sobie, a różnice dotyczące ciosów mogą wynikać z dymorfizmu płciowego. Tak więc pomysł ten nie jest już dziś akceptowany[4]. Stada Paraceratherium mogły migrować, żerując na wysokich drzewach, których nie mogli dosięgnąć mniejsi roślinożercy[26]. Osborn zasugerował, że sposób żerowania tego zwierzęcia przypominał raczej obserwowany u żywiących się wysoką roślinnością żyraf czy okapi leśne, a nie współczesnych nosorożcowatych, trzymających głowy blisko ziemi[36].
Rozmieszczenie geograficzne i siedlisko
Szczątki przypisywane Paraceratherium pochodzą z wczesno- do późnooligoceńskich (34–23 milionów lat temu) formacji euroazjatyckich leżących dziś na terenie Chin, Mongolii, Indii, Pakistanu, Kazachstanu, Gruzji, Turcji, Rumunii, Bułgarii oraz byłej Jugosławii[5]. Zasięg występowania rodzaju może korelować z paleogeograficznym rozwojem systemu górskiego alpejsko-himalajskiego. Zasięg znajdowanych pozostałości wskazuje, że zwierzęta te zamieszkiwały ciągłe masy lądowe tworzące podobne środowiska. Przeczą temu jednak mapy paleogeograficzne, które lokują tam liczne bariery morskie. Gatunek odniósł więc duży sukces, rozprzestrzeniając się wbrew nim[44]. Fauna żyjąca obok Paraceratherium obejmuje inne Rhinocerotoidea, parzystokopytne, gryzonie, amficjony, łasicowate, hienodonty, nimrawidy i kotowate[26].

Siedlisko Paraceratherium wydaje się zmieniać w obrębie jego zasięgu występowania. Wskazują na to typy formacji geologicznych, w których znajdowano ich szczątki[26]. Formacja Hsanda Gol leżąca w Mongolii reprezentuje suchy basen pustynny. Uważa się, że środowisko to porastały nieliczne wysokie drzewa i ograniczona pokrywa buszu, fauna zaś składała się głównie ze zwierząt żerujących na roślinności od czubków drzew do poziomu gruntu[45]. Badanie skamieniałych pyłków wykazało, że większość terenów dzisiejszych Chin porastał busz. Tworzyły go takie rośliny, jak łoboda, przęślowate i łużnik, wszystkie zaadaptowane do suchych siedlisk. Drzewa zdarzały się rzadko, skupiając się w pobliżu wód gruntowych[46]. Regiony Chin, na których żyły Paraceratherium, mieściły wyschnięte jeziora, obfitowały w wydmy. Najpopularniejsze skamieniałości roślinne tworzą tu liście zaadaptowanej do pustyń Palibinia. Drzewa Mongolii i Chin obejmują brzozy, wiązy, dęby i inne drzewa liściaste, podczas gdy na Syberii i w Kazachstanie występowały także orzechy włoskie[26]. Dera Bugti w Pakistanie porastała suchym lasem, umiarkowanym do podzwrotnikowego[42].
Powody, dla których Paraceratherium wyginęło po przetrwaniu około 11 milionów lat, pozostają nieznane. Wydaje się jednak nieprawdopodobne, by spowodowała to pojedyncza przyczyna[26]. Proponuje się tutaj zmianę klimatu, niskie tempo reprodukcji oraz inwazję należących do trąbowców gomfoterów z Afryki w późnym oligocenie. Gomfotery byłyby zdolne do wywołania znacznej zmiany podbijanych przez siebie siedlisk, podobnie jak dzisiaj czyni to słoń afrykański, niszczący drzewa i przekształcający lasy w tereny trawiaste. W końcu zasoby pokarmowe uległy znacznemu ograniczeniu i liczba Paraceratherium obniżyła się wraz z nimi, co uwrażliwiło populację na inne zagrożenia[47]. Duże drapieżniki, jak Hyaenaelurus czy Amphicyon, także przedostały się z Afryki do Azji, co odbyło się we wczesnym miocenie. Mogły one polować na młode Paraceratherium. W czasie tym pojawili się w Azji również inni roślinożercy[26].
Przypisy
- 1 2 3 4 Prothero 2013 ↓, s. 17–34.
- 1 2 3 C. Manias. Building Baluchitherium and Indricotherium: Imperial and International Networks in Early-Twentieth Century Paleontology. „Journal of the History of Biology”, 2014. DOI: 10.1007/s10739-014-9395-y. (ang.).
- 1 2 3 4 5 6 7 8 Prothero 2013 ↓, s. 67–86.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 Prothero 2013 ↓, s. 87–106.
- 1 2 3 Prothero 2013 ↓, s. 35–52.
- ↑ G.E. Pilgrim. Notices of new mammalian genera and species from the Tertiaries of India. „Records of the Geological Survey of India”. 40 (1), s. 63–71, 1910.
- 1 2 C.F. Cooper. On the Skull and Dentition of Paraceratherium bugtiense: A Genus of Aberrant Rhinoceroses from the Lower Miocene Deposits of Dera Bugti. „Philosophical Transactions of the Royal Society B: Biological Sciences”. 212 (391–401), s. 369–394, 1924. DOI: 10.1098/rstb.1924.0009. (ang.).
- ↑ C. Forster-Cooper. LXXVIII. – Paraceratherium bugtiense, a new genus of Rhinocerotidae from the Bugti Hills of Baluchistan.–Preliminary notice. „Annals and Magazine of Natural History Series 8”. 8 (48), s. 711–716, 1911. DOI: 10.1080/00222931108693085. (ang.).
- ↑ C. Forster-Cooper. XLIV.– Thaumastotherium osborni, a new genus of perissodactyles from the Upper Oligocene deposits of the Bugti hills of Baluchistan. –Preliminary notice. „Annals and Magazine of Natural History Series 8”. 12 (70), s. 376–381, 1913. DOI: 10.1080/00222931308693412. (ang.).
- ↑ C. Forster-Cooper. Correction of generic name. „Annals and Magazine of Natural History Series 8”. 12 (71), s. 504, 1913. DOI: 10.1080/00222931308693431. (ang.).
- ↑ C. Forster-Cooper. Baluchitherium osborni (? syn. Indricotherium turgaicum, Borrissyak). „Philosophical Transactions of the Royal Society of London”. 212, s. 35–66, 1923. DOI: 10.1098/rstb.1924.0002. JSTOR: 92060.
- ↑ M. Pavlova. Indricotherium transouralicum n. sp. provenant du district de Tourgay. „Bulletin de la Societe des Naturalistes de Moscou, Section Geologique”. 31, s. 95–116, 1922. (fr.).
- ↑ A.A. Borissiak. Über die Unterfamilie Indricotheriinae Boriss. = Baluchitheriinae Osb. „Zentralblatt für Mineralogie, Geologie und Paläontologie”. 18, s. 571–575, 1924. (niem.).
- 1 2 3 4 5 6 7 8 9 S.G. Lucas, J.C. Sobus: The Systematics of Indricotheres. W: D.R. Prothero, R.M. Schoch: The Evolution of Perissodactyls. New York, New York & Oxford, England: Oxford University Press, 1989, s. 358–378. ISBN 978-0-19-506039-3. OCLC 19268080.
- ↑ Prothero 2013 ↓, s. 1–16.
- ↑ H.F. Osborn, The extinct giant rhinoceros ''Baluchitherium'' of Western and Central Asia, „Natural History”, 23, 1923 (3), s. 208–228 (ang.).
- ↑ Q. Zhan-Xiang, A new genus of giant rhinoceros from oligocene of Dzungaria, Sinkang, „Vertebrata Palasiatica”, 2, 1973 (11), s. 182–191 (chiń.).
- 1 2 3 W. Granger, W.K. Gregory. Further notes on the gigantic extinct rhinoceros, Baluchitherium, from the Oligocene of Mongolia. „Bulletin of the American Museum of Natural History”. 72, s. 1–73, 1936.
- ↑ C. Forster-Cooper. The Extinct Rhinoceroses of Baluchistan. „Philosophical Transactions of the Royal Society B: Biological Sciences”. 223 (494–508), s. 569–616, 1934. DOI: 10.1098/rstb.1934.0013. (ang.).
- ↑ P.O. Antoine, L. Karadenizli, G.E. Saraç, S. Sen. A giant rhinocerotoid (Mammalia, Perissodactyla) from the Late Oligocene of north-central Anatolia (Turkey). „Zoological Journal of the Linnean Society”. 152 (3), s. 581–592, 2008. DOI: 10.1111/j.1096-3642.2007.00366.x. (ang.).
- ↑ Y. Ye, Meng J., W.W. Yu. Discovery of Paraceratherium in the northern Junggar Basin of Xinjiang. „Vertebrata Palasiatica”. 41, s. 220–229, 2003. (chiń. • ang.).
- ↑ Zhan-Xiang Qui, Ban-Yue Wang, Tao Deng. Indricotheres (Perissodactyla, Mammalia) from Oligocene in Linxia Basin, Gansu, China. „Vertebrata Palasiatica”. 42 (3), s. 177–192, 2004. (ang.).
- 1 2 3 4 Prothero 2013 ↓, s. 53–66.
- ↑ L.B. Radinsky. The Families of the Rhinocerotoidea (Mammalia, Perissodactyla). „Journal of Mammalogy”. 47 (4), s. 631, 1966. DOI: 10.2307/1377893. (ang.).
- ↑ L. Holbrook. The Phylogeny and Classification of Tapiromorph Perissodactyls (Mammalia). „Cladistics”. 15 (3), s. 331–350, 1999. DOI: 10.1006/clad.1999.0107. (ang.).
- 1 2 3 4 5 6 7 8 9 Prothero 2013 ↓, s. 107–121.
- ↑ S.G. Lucas, R.M. Schoch, E. Manning. The Systematics of Forstercooperia, a Middle to Late Eocene Hyracodontid (Perissodactyla: Rhinocerotoidea) from Asia and Western North America. „Journal of Paleontology”. 55 (4), s. 826–841, 1981. JSTOR: 1304430.
- 1 2 3 M. Fortelius, J. Kappelman. The largest land mammal ever imagined. „Zoological Journal of the Linnean Society”. 108, s. 85–101, 1993. DOI: 10.1111/j.1096-3642.1993.tb02560.x. (ang.).
- ↑ T. Tsubamoto. Estimating body mass from the astragalus in mammals. „Acta Palaeontologica Polonica”, s. 259–265, 2012. DOI: 10.4202/app.2011.0067. (ang.).
- ↑ A. Larramendi. Shoulder height, body mass and shape of proboscideans. „Acta Palaeontologica Polonica”, 2015. DOI: 10.4202/app.00136.2014. (ang.).
- ↑ G.S. Paul. Dinosaur models: The good, the bad, and using them to estimate the mass of dinosaurs. „Dinofest International Proceedings”, s. 129–142, 1997.
- ↑ M.P. Taylor, M.J. Wedel. Why sauropods had long necks; and why giraffes have short necks. „PeerJ”. 1, s. e36, 2013. DOI: 10.7717/peerj.36. PMID: 23638372. (ang.).
- ↑ P.M. Sander, A. Christian, M. Clauss, R. Fechner i inni. Biology of the sauropod dinosaurs: The evolution of gigantism. „Biological Reviews”. 86 (1), s. 117–155, 2011. DOI: 10.1111/j.1469-185X.2010.00137.x. PMID: 21251189. (ang.).
- 1 2 3 4 P.O. Antoine, S.M. Ibrahim Shah, I.U. Cheema, J.Y. Crochet i inni. New remains of the baluchithere Paraceratherium bugtiense from the Late/latest Oligocene of the Bugti hills, Balochistan, Pakistan. „Journal of Asian Earth Sciences”. 24, s. 71–77, 2004. DOI: 10.1016/j.jaes.2003.09.005. (ang.).
- ↑ W. Granger, W.K. Gregory. A revised restoration of the skeleton of Baluchitherium, gigantic fossil rhinoceros of Central Asia. „American Museum novitates”. 787, s. 1–3, 1935.
- 1 2 H.F. Osborn. Baluchitherium grangeri, a giant hornless rhinoceros from Mongolia. „American Museum novitates”. 78, s. 1–15, 1923.
- ↑ V.L. Gromova. Gigantskie nosorogi. „Trudy Paleontology Institut Akademii Nauk SSSR”. 71, s. 154–156, 1959. (ros.).
- ↑ P.O. Antoine. There were giants upon the earth in those days (Book Review). „Palaeovertebrata”. 38, s. 1–3, 2014. (ang.).
- ↑ Naish, D: Tet Zoo Bookshelf: van Grouw’s Unfeathered Bird, Bodio’s Eternity of Eagles, Witton’s Pterosaurs, Van Duzer’s Sea Monsters on Medieval and Renaissance Maps!. [w:] blogs.scientificamerican.com/tetrapod-zoology [on-line]. 2013-06-29. [dostęp 2014-10-05].
- ↑ R.M. Alexander. All-time giants: the largest animals and their problems. „Palaeontology”. 41 (6), s. 1231–1245, 1998.
- ↑ M. Clauss, R. Frey, B. Kiefer, M. Lechner-Doll i inni. The maximum attainable body size of herbivorous mammals: Morphophysiological constraints on foregut, and adaptations of hindgut fermenters. „Oecologia”. 136 (1), s. 14–27, 2003. DOI: 10.1007/s00442-003-1254-z. PMID: 12712314. (ang.).
- 1 2 C. Martin, I. Bentaleb, P. -O. Antoine. Pakistan mammal tooth stable isotopes show paleoclimatic and paleoenvironmental changes since the early Oligocene. „Palaeogeography, Palaeoclimatology, Palaeoecology”. 311, s. 19–29, 2011. DOI: 10.1016/j.palaeo.2011.07.010. (ang.).
- ↑ Y. Wang, T. Deng. A 25 m.y. Isotopic record of paleodiet and environmental change from fossil mammals and paleosols from the NE margin of the Tibetan Plateau. „Earth and Planetary Science Letters”. 236, s. 322–338, 2005. DOI: 10.1016/j.epsl.2005.05.006. (ang.).
- ↑ S. Sen, P.O. Antoine, B. Varol, T. Ayyildiz i inni. Giant rhinoceros Paraceratherium and other vertebrates from Oligocene and middle Miocene deposits of the Kağızman-Tuzluca Basin, Eastern Turkey. „Naturwissenschaften”. 98 (5), s. 407–423, 2011. DOI: 10.1007/s00114-011-0786-z. PMID: 21465174. (ang.).
- ↑ J. Meng, M.C. McKenna. Faunal turnovers of Palaeogene mammals from the Mongolian Plateau. „Nature”. 394 (6691), s. 364–367, 1998. DOI: 10.1038/28603. (ang.).
- ↑ E. B. Leopold, G. Liu, S. Clay-Poole: Low-Biomass Vegetation in the Oligocene?. W: D.R. Prothero, W.A. Berggren: Eocene-Oligocene Climatic and Biotic Evolution. Princeton: Princeton University Press, 1992, s. 399–420. ISBN 978-0-691-02542-1.
- ↑ P.V. Putshkov. „Proboscidean agent” of some Tertiary megafaunal extinctions. „Terra degli elefanti Congresso internazionale: The world of elephants”, s. 133–136, 2001.
Bibliografia
- D. Prothero: Rhinoceros Giants: The Palaeobiology of Indricotheres. Indiana: Indiana University Press, 2013. ISBN 978-0-253-00819-0.