| Rhinocerotidae | |||||
| J.E. Gray, 1821[1] | |||||
.jpg.webp) Przedstawiciel rodziny – nosorożec biały (Ceratotherium simum) | |||||
| Systematyka | |||||
| Domena | |||||
|---|---|---|---|---|---|
| Królestwo | |||||
| Typ | |||||
| Podtyp | |||||
| Gromada | |||||
| Podgromada | |||||
| Infragromada | |||||
| Rząd | |||||
| Podrząd | |||||
| Rodzina |
nosorożcowate | ||||
| Typ nomenklatoryczny | |||||
|
Rhinoceros Linnaeus, 1758 | |||||
| |||||
| Rodzaje | |||||
| |||||
| Zasięg występowania | |||||
 | |||||
Nosorożcowate[59], nosorożce (Rhinocerotidae) – rodzina dużych ssaków z rzędu nieparzystokopytnych (Perissodactyla). Kiedyś liczyła 70 rodzajów, z których obecnie żyją 4 liczące 5 gatunków, wszystkie zagrożone wyginięciem, z czego trzy krytycznie zagrożone wyginięciem, w tym nosorożec jawajski uznawany za najbardziej zagrożonego ssaka świata. Osiągają dość duże rozmiary i masę ciała, największy jest nosorożec biały. Ich cechą charakterystyczną jest keratynowy róg bądź dwa rogi na nosie, większe u gatunków afrykańskich, osiągające 130 cm. Zamieszkują różnorodne siedliska, od suchych do podmokłych, żywiąc się trawą bądź wyższą roślinnością. Samce wykazują pewne cechy terytorialne i brutalnie walczą o samicę, wykorzystując rogi bądź ciosy. Samica rodzi zwykle jedno młode po długiej ciąży, w którą zachodzi co 2-3 lata. Intensywne polowania celem zdobycia rogów znacznie zmniejszyły zasięg występowania i liczebność wszystkich gatunków.
Genetyka
Większość nosorożcowatych cechuje się diploidalną liczbą chromosomów 82, aczkolwiek nosorożec czarny ma 84. Jest to prawie największa liczba chromosomów wśród ssaków, jedynie pewne gatunki gryzoni mają ich więcej w genomie. Ponadto w kariotypie nosorożca białego południowego dostrzeżono translokację robertsonowską[60].
Budowa

Nosorożcowate należą do największych współczesnych ssaków, mimo że nie dorastają rozmiarów swych wymarłych krewnych Indricotheriinae. Obecnie największy przedstawiciel rodziny, nosorożec biały, jest czwartym największym lądowym ssakiem. Wyprzedzają go jedynie 3 gatunki słoni. Waży pomiędzy 1350 a 3500 kg. Kolejne, piąte, miejsce zajmuje nosorożec czarny. Waży między 800 a 1300 kg. Z drugiej strony najmniejszym przedstawiciel nosorożcowatych jest obecnie nosorożec sumatrzański o masie ciała między 600 a 950 kg. Niemniej w przeciwieństwie do nosorożców z Afryki i nosorożca indyjskiego nosorożce sumatrzański i jawajski poznane są słabo i nawet podstawowe dane o ich wielkości czy masie ciała są nieliczne[60].
Długość ciała 236–420 cm, długość ogona 25–70 cm, wysokość w kłębie 100–200 cm; masa ciała 600–3500 kg[60].
Dymorfizm płciowy raczej nie występuje, samce i samice są zwykle podobnej wielkości bądź samce nieznacznie tylko przerastają płeć przeciwną. W przypadku nosorożców indyjskich hodowanych w niewoli samce są istotnie większe od samic, czego nie obserwuje się a naturze[60].
Najbardziej zwracającą uwagę cechą dzisiejszych nosorożcowatych, od której zresztą bierze źródłosłów nazwa rodziny, jest obecność rogów. U afrykańskich przedstawicieli rodziny występują zwykle 2 rogi, większy nosowy i mniejszy czołowy. Dwa znacznie mniejsze rogi zdobią też głowę nosorożca sumatrzańskiego. Przedstawiciele rodzaju typowego mają tylko jeden róg. U nosorożca czarnego róg nosowy może osiągać aż 130 cm długości. Znacznie mniej, od 2 do 55 cm, mierzy róg czołowy. Nosorożec biały nosi róg nosowy mający od 94 do 102 cm i róg czołowy osiągający także do 55 cm. Najmniejsze rogi ma prawdopodobnie nosorożec sumatrzański. U okazów muzealnych tego gatunku mierzą one od 25 do 80 cm, ale trudno powiedzieć, czy wielkość ta jest reprezentatywna dla dziko żyjącej populacji. Nosorożec indyjski ma róg wielkości 24-25 cm, podobny róg występuje u nosorożca jawajskiego. W przeciwieństwie do wielu innych rogatych zwierząt, jak wołowate, kameleony czy inne jaszczurki, rogi nosorożców nie posiadają kostnego rdzenia. W całości buduje je keratyna ze zmagazynowanym wapniem i melaniną. Podobną budowę mają paznokcie innych zwierząt. Czyni to rogi nosorożców podatnym na zużycie. Ścierają się one z czasem. Z drugiej strony cały czas rosną, nawet aż 7 cm na rok, jak u nosorożca białego, jednakże wyjściowy efekt często sumuje się od zera. Ścieranie natomiast czyni rogi cieńszymi u czubka. Ponadto skład chemiczny rogu zależy od miejsca życia osobnika. Obserwowano bezrogie samice. Także pierwsze nosorożcowate w zapisie kopalnym nie miały jeszcze rogów, pojawiły się one później, pierwszy raz u oligoceńskiego Diceratherium[60].
Funkcją rogów jest ochrona zwierzęcia. Zaniepokojony nosorożec czarny szarżuje przed siebie, aczkolwiek jego szarża rzadko przeradza się w rzeczywisty atak. Rodzi to pytania o zasadność odcinania rogów zwierzętom celem ochrony ich przed kłusownikami. Istnieje bowiem obawa, że pozbawienie matki rogu, a więc narzędzia obrony, znacząco zwiększa śmiertelność młodych padających ofiarą lwów i krokut. Samce nosorożców używają rogów także w walkach między sobą[60].
Taksony azjatyckie odróżniają się od afrykańskich obecnością ciosów w żuchwie samców, osiągających u nosorożca indyjskiego aż 9,5 cm, u pozostałych dwóch gatunków znacznie mniej. Są ostre. Ich również samce używają w walkach z innymi samcami, ale też w stosunku do samic i cieląt. Nosorożce azjatyckie w mniejszym stopniu walczą za pomocą rogów, pozostających jedyną bronią ich afrykańskich krewnych. Prawdopodobnie gruba i pofałdowana skóra otaczająca okolice karku i ramion tych zwierząt stanowi rodzaj ochrony przed ugryzieniami innych samców[60].
Charakterystyczną cechą nosorożca czarnego jest długa, chwyta górna warga, ułatwiająca mu zrywanie wyższej roślinności. Natomiast nosorożec biały posiada szerokie wargi przydatne w pasieniu się trawą. Od tejże wargi pochodzi prawdopodobnie jego nazwa, jako że nie jest on barwy białej. Holenderskie wijd stanowiło odpowiednik angielskiego wide, szeroki, nie white, biały, i odnosiło się do szerokiej wargi właśnie[60].
Wzrok jest słaby i nosorożce słabą widzą na odległość. Mierzące się wzrokiem samce muszą zbliżyć się do siebie na kilka metrów, by się widzieć. Znacznie lepiej rozwinęły się słuch i węch tych ssaków, ten ostatni wykorzystywany do znakowania terytoriów odchodami i moczem[60].
Kończyny wydają się krótkie, niemniej na krótkich dystansach umożliwiają zwierzęciu osiągnięcie prędkości 55 km/h. Kończą się trzema palcami. U zamieszkujących pustynie nosorożców czarnych istotną rolę odgrywają mięsiste poduszki pod kopytami, pomagające utrzymać stabilność na piasku[60].
Systematyka

Nosorożcowate są rodziną należącą do rzędu nieparzystokopytnych[60]. W dziewiętnastowiecznych ujęciach współtworzyły rząd gruboskórych[61].
Do rodziny należą następujące występujące współcześnie rodzaje[62][63][64][59]:
- Ceratotherium J.E. Gray, 1868 – jedynym żyjącym współcześnie przedstawicielem jest Ceratotherium simum (Burchell, 1817) – nosorożec biały
- Diceros J.E. Gray, 1821 – jedynym żyjącym współcześnie przedstawicielem jest Diceros bicornis (Linnaeus, 1758) – nosorożec czarny
- Rhinoceros Linnaeus, 1758
- Dicerorhinus Gloger, 1841 – jedynym występującym współcześnie przedstawicielem jest Dicerorhinus sumatrensis (G. Fischer, 1814) – nosorożec sumatrzański
Opisano również szereg rodzajów wymarłych zgrupowanych w zależności od ujęcia systematycznego w różnych podrodzinach i plemionach (tutaj nie wyróżnianych)[65]:
- Aceratherium Kaup, 1832[66]
- Acerorhinus Kretzoi, 1942[67]
- Alicornops Ginsburg & Guérin, 1979[68]
- Amphicaenopus Wood, 1927[69] – jedynym przedstawicielem był Amphicaenopus platycephalus (Osborn & Wortman, 1894)
- Aphelops Cope, 1873[70]
- Aprotodon Forster Cooper, 1915[71]
- Brachypotherium Roger, 1904[72]
- Bugtirhinus Antoine & Welcomme, 2000[73] – jedynym przedstawicielem był Bugtirhinus praecursor Antoine & Welcomme, 2000
- Caementodon Heißig, 1972[74]
- Chilotheridium Hooijer, 1971[75]
- Chilotherium Ringström, 1924[76]
- Coelodonta Bronn, 1831[77]
- Diaceratherium Dietrich, 1931[78]
- Diceratherium Marsh, 1875[79]
- Dihoplus Brandt, 1878[80]
- Dromoceratherium Crusafont i Pairó, Villalta-Comella & Truyols Santonja, 1955[81] – jedynym przedstawicielem był Dromoceratherium mirallesi Crusafont i Pairó, Villalta Comella & Truyols, 1955
- Elasmotherium G. Fischer, 1808[82] – elasmoterium
- Eoazara Geraads & Zouhri, 2021[83] – jedynym przedstawicielem był Eoazara xerii Geraads & Zouhri, 2021
- Epiaceratherium Abel, 1910[84]
- Floridaceras Wood, 1964[85] – jedynym przedstawicielem był Floridaceras whitei Wood, 1964
- Gaindatherium Colbert, 1934[86]
- Galushaceras Prothero, 2005[87] – jedynym przedstawicielem był Galushaceras levellorum Prothero, 2005
- Gulfoceras Albright, 1999[88] – jedynym przedstawicielem był Gulfoceras westfalli Albright, 1999
- Hispanotherium Crusafont i Pairó & Villalta Comella, 1947[89]
- Hoploaceratherium Ginsburg & Heißig, 1989[90]
- Huaqingtherium Huang Wanpo & Yan Defa, 1983[91]
- Iranotherium Ringström, 1924[92] – jedynym przedstawicielem był Iranotherium morgani (Mecquenem, 1908)
- Itanzatherium Belyayeva, 1966[93] – jedynym przedstawicielem był Itanzatherium angustirostre Belyayeva, 1966
- Kenyatherium Aguirre & Guérin, 1974[94] – jedynym przedstawicielem był Kenyatherium bishopi Aguirre & Guérin, 1974
- Lartetotherium Ginsburg, 1974[95] – jedynym przedstawicielem był Lartetotherium sansaniense (Lartet, 1848)
- Menoceras Troxell, 1921[96]
- Mesaceratherium Heißig, 1969[97]
- Miodiceros Giaourtsakis, 2022[98] – jedynym przedstawicielem był Miodiceros neumayri (Osborn, 1900)
- Molassitherium Becker & Antoine, 2013[99]
- Nesorhinus Antoine, Reyes, Amano, Bautista, Chang, Claude, De Vos & Ingicco, 2022[100]
- Ningxiatherium Chen Guanfang, 1977[101]
- Ougandatherium Guérin & Pickford, 2003[102] – jedynym przedstawicielem był Ougandatherium napakense Guérin & Pickford, 2003
- Paradiceros Hooijer, 1968[103] – jedynym przedstawicielem był Paradiceros mukirii Hooijer, 1968
- Penetrigonias Tanner & Martin, 1976[104]
- Peraceras Cope, 1880[105]
- Parelasmotherium Killgus, 1923[106]
- Persiatherium Pandolfi, 2015[107]
- Plesiaceratherium Yang Zhongjian, 1937[108]
- Pleuroceros Roger, 1898[109]
- Procoelodonta Matthew, 1931[110]
- Prohyracodon Koch, 1897[111]
- Prosantorhinus Heißig, 1974[112]
- Protaceratherium Abel, 1910[113]
- Ronzotherium Aymard, 1854[114]
- Rusingaceros Geraads, 2010[115] – jedynym przedstawicielem był Rusingaceros leakeyi (Hooijer, 1966)
- Samburuceros Handa, Nakatsukasa, Kunimatsu & Nakaya, 2017[116] – jedynym przedstawicielem był Samburuceros ishidai Handa, Nakatsukasa, Kunimatsu & Nakaya, 2017
- Shansirhinus Kretzoi, 1942[67]
- Shennongtherium Huang Wanpo & Yan Defa, 1983[117] – jedynym przedstawicielem był Shennongtherium hypsodontum Huang Wanpo & Yan Defa, 1983
- Sinotherium Ringström, 1923[118] – jedynym przedstawicielem był Sinotherium lagrelii Ringström, 1923
- Skinneroceras Prothero, 2005[119] – jedynym przedstawicielem był Skinneroceras manningi Prothero, 2005
- Stephanorhinus Kretzoi, 1942[120]
- Subchilotherium Heißig, 1975[121]
- Subhyracodon Brandt, 1878[122]
- Symphysorrhachis Belyayeva, 1954[123] – jedynym przedstawicielem był Symphysorrhachis brevirostris Belyayeva, 1954
- Teleoceras Hatcher, 1894[124]
- Teletaceras Hanson, 1989[125]
- Trigonias F.A. Lucas, 1900[126]
- Turkanatherium Deraniyagala, 1951[38] – jedynym przedstawicielem był †Turkanatherium acutirostratum Deraniyagala, 1951
- Uintaceras Holbrook & S.G. Lucas, 1997[127]
- Victoriaceros Geraads, McCrossin & Benefit, 2012[128]
- Woodoceras Prothero, 2005[129] – jedynym przedstawicielem był Woodoceras brachyops Prothero, 2005.
Ewolucja
_-_Mauricio_Ant%C3%B3n.jpg.webp)
Nosorożcowate są starą rodziną ssaków. Pojawiły się jeszcze w epoce eoceńskiej, wywodząc się z tej samej grupy, co dwie inne blisko spokrewnione rodziny nosorożców, Hyracodontidae, do których zaliczały się największe lądowe ssaki wszech czasów, jak Paraceratherium, a także prowadzące częściowo wodny tryb życia Amynodontidae. Pierwsze właściwe nosorożcowate pojawiły się w Euroazji i były niewielkimi zwierzętami, dopiero późnej zwiększając swe rozmiary zgodnie z zasadą Cope'a. Nie miały też jeszcze rogów. W oligocenie osiągnęły już znaczną różnorodność, choć nie opanowały jeszcze Afryki, z którą kojarzą się obecnie. W epoce tej istniały już formy o średnich i dużych rozmiarach. W oligocenie pojawiły się też pierwsze rogate nosorożce, Diceratherium. Był to także jedyny znany przedstawiciel amerykańskiej megafauny przez 10 milionów lat[60].
Nosorożcowate osiągnęły istotny sukces ewolucyjny, obejmując szeroki zakres nisz ekologicznych. Dzięki dużym rozmiarom mogły wykarmić się szorstkim, słabej jakości pokarmem, w tym niską roślinnością bogatą w krzemionkę, na co odpowiedziały molaryzacją przedtrzonowcow. Jej stopień nie był jednak w obrębie gatunku cechą stałą, co doprowadziło do istotnego zamieszania w XX-wiecznej paleontologii, kiedy to na podstawie takich niestałych cech opisywano taksony, które potem okazywały się synonimiczne. Zwierzęta te zasiedliły Amerykę, Europę i Azję, a także Afrykę. Ich sukces z oligocenu nie trwał jednak wiecznie, jako że w miocenie doszło do wymarcia części linii w skutek zmian klimatycznych. Wymarły prawie wszystkie nosorożcowate amerykańskie, w Eurazji przetrwały tylko dwie linie. Przedstawiciele jednej z nich przypominały dzisiejsze nosorożce sumatrzańskie. Z tej właśnie linii wywodzi się sławny nosorożec włochaty, zamieszkujący w plejstocenie tereny północy Euroazji. Do dzisiaj nosorożce sumatrzańskie cechują się owłosieniem większym, niż pozostałe nosorożcowate, zwłaszcza młode osobniki, tracące włosy z biegiem życia. Z drugiej strony pozycja nosorożca sumatrzańskiego na drzewie rodowym nosorożcowatych nie została do końca wyjaśniona. Zazwyczaj zalicza się go do azjatyckiej podrodziny Rhinocerotinae, wraz z rodzajem Rhinoceros, obejmującym nosorożce indyjskiego i jawajskiego. Jednakże część autorów uważa go za bliższego nosorożcowatym afrykańskim, z którymi łączy go posiadanie obu rogów[60].
W przeciwieństwie do starego nosorożca sumatrzańskiego gatunki z rodzaju Rhinoceros są znacznie młodsze. Nosorożec indyjski i nosorożec jawajski pojawiły się w środkowym plejstocenie[60].
W Afryce żyją obecnie prawdopodobnie dwa gatunki nosorożcowatych, nosorożec czarny i nosorożec biały. Ten ostatni obejmuje dwa podgatunki (nosorożec północny i południowy), których zasięg rozdziela prawdopodobnie rzeka Zambezi. W przeciwieństwie do świetnie pływających nosorożców Azji nosorożce białe nie należą do dobrych pływaków i nie potrafią przekroczyć większej rzeki. Oba podgatunki nie kontaktują się ze sobą prawdopodobnie od miliona lat, co skłoniło część autorów do uznania ich za osobne gatunki. Kontestatorzy tego pomysłu zwracają jednak uwagę na niewielkie różnice morfologiczne między nosorożcem północnym i południowym, optując za pozostawieniem obu form jako podgatunków[60].
Tryb życia
Cykl życiowy

Nosorożcowate rozmnażają się powoli. Samice zaczynają miesiączkować i być zdolne do zapłodnienia już w wieku około 3 (nosorożec indyjski) czy 3,7 lat (nosorożec biały), co obserwowano w niewoli. Na wolności, gdzie warunki pokarmowe są znacznie gorsze, rzadko jednak dochodzi do takich sytuacji, a trzyletnie źrebię może jeszcze przebywać pod matczyną opieką. Przedłużanie gatunku zaczyna się raczej w wieku 6-7 lat, przynajmniej w przypadku nosorożców białego, czarnego i indyjskiego, jako że dane dotyczące nosorożców sumatrzańskiego i jawajskiego są ubogie. Jeszcze później do rozrodu przystępują samce, w wieku 13-15 lat. Osiągnięcie dojrzałości pod względem anatomicznym nie oznacza bowiem wcale, że samiec może spokojnie przystąpić do przekazywania swych genów następnemu pokoleniu. Nie chodzi tylko o rzadkie występowanie samic zdolnych do rozrodu, dojrzewających późno i odbywających długie przerwy pomiędzy kolejnymi ciążami. Na obszarami, gdzie zbiera się dużo płodnych samic, obecne mogą też być starsze, większe samce, blokujące swym młodszym konkurentom dostęp do samic. Nosorożce potrafią walczyć bardzo brutalnie. Nosorożce indyjskie używają w tym celu bardziej ciosów żuchwy niż rogów, co potwierdzają obserwacje samców o ułamanym rogu i wysokim statusie. To raczej utrata ciosów może się dla samca skończyć źle, nawet zgonem. Ciosy powoli odrastają, jednak nim odrosną, samiec nie jest w stanie obronić się przed atakami. Samce kąsają konkurentów, zwłaszcza w okolice głowy i ramion, pokrytych grubą, zasobną w fałdy skórą[60].
Agresywne zachowania samców skutkować mogą zgonem nawet połowy z nich, ale też 30% samic. Samica w rui może przemierzyć terytoria kilku samców, które interesują się jej stanem, sprawdzając jej mocz. Zaloty mogą trwać od 5 do 20 dni, przebiegają również bardzo agresywnie, obejmują też długie pościgi. U wszystkich nieparzystokopytnych zaloty mają w sobie dużo agresji, jednak u nosorożcowatych zjawisko to występuje intensywniej. Jego funkcja rodzi pytania. Samiec krzywdzący jedyną płodną samicę w pobliżu nie osiągnie bowiem dzięki temu większego sukcesu rozrodczego. Być może zachowania takie służą zmuszeniu do kopulacji samic niechętnych rozmnażaniu, na przykład wobec dostępności większych samców. Agresja dorosłych samców może także przejawiać się poprzez zabijanie młodych spłodzonych przez inne osobniki. Samica wtedy szybko wchodzi w ruję, umożliwiając agresorowi spłodzenie potomka. Kopulacja trwa od pół do całej godziny. Pod koniec samica daje znać samcowi, wychodzi spod niego. On jednak jeszcze jakiś czas spędza w jej otoczeniu, pilnując, by nie zbliżył się do niej inny samiec[60].
Ciąża trwa długo, u nosorożca indyjskiego 16 miesięcy, podobnie u nosorożca białego. Najczęściej nie występuje wyraźny okres narodzin, aczkolwiek u nosorożca czarnego występują dwa piki (styczeń-luty i czerwiec-sierpień), podobnie u białego (marzec i lipiec). Pomiędzy kolejnymi porodami upływa 2-2,5 lat. Córka nosorożca białego może po kolejnym porodzie jej matki powtórnie do niej dołączyć. Natomiast samica pozostaje płodna długo. Obserwowano samice ponadczterdziestoletnie, które w dalszym ciągu rodziły. Samice potrafią dożyć 50 lat, prowadzące bardziej gwałtowne życie samce dożywają 30 lat. Zazwyczaj rodzi się pojedyncze młode, rzadko zdarzają się bliźnięta. Jak tylko wstanie, noworodek poszukuje sutków matki, zazwyczaj już 2-3 godziny po porodzie. Przez pierwsze dni chwieje się na nogach. Młode szybko rośnie, przybierając u nosorożca indyjskiego 2-3 kg dziennie w niewoli. Cielęta żyjące na wolności są mniejsze, nawet o połowę. Zważone martwe czteromiesięczne młode ważyło przykładowo 158 kg. Karmienie mlekiem trwa u nosorożca białego rok, niekiedy półtora, przy czym samce ssą dłużej od samic. Matka może zajmować się młodym kilka lat. Niekiedy pozostawia młode, np. kiedy zbliża się do niej inny samiec, ale potem do niego wraca[60].
Struktura społeczna

Nosorożcowate zazwyczaj żyją samotnie. Samce mogą przejawiać zachowania terytorialne. Oznaczają swe terytorium moczem, który potrafią rozpylać na odległość 5 m, i odchodami, które rozgrzebują kończynami bądź też zdobią rozgrzebaną uprzednio ziemię. Korzystają też ze wspólnych latryn, oceniając dzięki nim węchem, które osobniki znajdują się w pobliżu. Zazwyczaj tolerują znane sobie zwierzęta. Natomiast w stosunku do nowych, nieznanych sobie osobników reagują agresją, która może skutkować zgonem. Szacuje się, że połowa samców nosorożca czarnego i 30% samic odchodzi na skutek ran zadanych im przez inne osobniki tego samego gatunku, i to pomimo ogólnej wysokiej odporności nosorożców, dobrze znoszących urazy. Niegdyś 20 nosorożców czarnych przeniesiono do Parku Narodowego Etosha, zamieszkiwanego już przed 11 osobników. W ciągu kolejnych sześciu tygodni poległo 5 z nowych zwierząt. Walkę nosorożce rozpoczynają od zmierzenia się wzrokiem, a że wzrok mają słaby, muszą to robić z bliska. Opuszczają głowy i szorują rogiem po ziemi. Nosorożce azjatyckie także prezentują ciosy i porykują. Sikają na odległość. Potem krzyżują rogi. Taksony azjatyckie używają także ciosów, zadając rany. W końcu jeden z walczących ssaków wycofuje się i ucieka. Wygrany często goni go, starając się gryźć po szyi, bokach, zadzie i brzuchu. Nosorożec indyjski często potem potrafi poszukiwać konkurenta. Jeśli przegrany samiec przeżyje, co nie jest oczywiste, wycofuje się. Niekiedy osiada na obszarze przyległym do granic terytorium zwycięskiego konkurenta, niekiedy pozostaje na części swych dawnych terenów, nie ośmielając się dalej oznaczać ich, co od tej pory będzie czynił zwycięski samiec. Nosorożce noszą blizny po walkach, świadczące o ranach, których nie przeżyłyby inne ssaki[60].
Samice mogą żyć samotnie, mogą też zbierać się w grupy. U nosorożca czarnego jest do rzadkie, aczkolwiek niekiedy mała liczba samic wspólnie żeruje. Córka może pozostać pod opieką matki po urodzeniu przez nią kolejnego dziecka. Znacznie częściej zbierają się samice nosorożca białego, niekiedy na raz kilkanaście sztuk. Obserwowano razem kilkanaście zwierząt, w tym samice z młodymi. Obserwowano też ponad 30 nosorożców indyjskich w jednym zbiorniku wodnym[60].
Ssaki te komunikują się ze sobą za pomocą węchu bądź słuchu, jako że wzrokiem dysponują słabym. Wydają z siebie różnorodne odgłosy. W przypadku nosorożca indyjskiego naliczono ich 10. Jednym z najczęstszych jest parsknięcie, towarzyszące spotkaniom zwierząt tego samego gatunku, najczęściej wydawane 2 razy, z przedziałem od jednego do dwudziestu. Do najgłośniejszych należy trąbnięcie (honk), często wydawane przez pokonane osobniki w walkach czy podczas ucieczki przed dominantem, niosące się na odległość kilometrów. Poddające się osobniki także beczą. Do innych głosów należą ryk oraz pisk, ten ostatni towarzyszący samcom zalecającym się do płci przeciwnej. W relacji matki i cielęcia słyszy się natomiast cichsze muczenie, niesłyszalne z większych odległości. Nosorożec czarny również parska na powitanie, a podczas walk wydaje z siebie trąbiące dźwięki. Gniew wyraża także parskaniem i odgłosami przypominającymi kichanie. Niepokój przekazuje dźwiękiem "wonk", przechodzącym w krzyk, podczas gdy "mmwonk" wiąże się z odczuwanym spokojem. Szybszy oddech pojawiać się może zarówno w razie niepokoju, uspokojenia, jak i na powitanie innego osobnika[60].
Samice raczej nie wykazują terytorializmu, areały sąsiadujących samic zwykle pokrywają się w istotnym stopniu ze sobą. Samce również nie są w pełni terytorialne, ale w czasach grupowania się płodnych samic wykazują pewne zachowania terytorialne, a nawet walczą do śmierci. Nosorożce czarne mogą też bronić dostępu do terenów dostarczających atrakcyjnego pokarmu. Nosorożce indyjskie większość czasu spędzają wzdłuż rzek, rzadko oddalając się od nich na dalej niż 2 km. Nosorożce białe dziennie przemierzają dziennie 5 km w porze deszczowej. Podczas pory suchej przemierzają większe odległości, jako że rosną odległości do najbliższych zbiorników wodnych. Nosorożce poruszają się zygzakiem, zakreślając pętle. Powierzchnie zajmowane przez nosorożce wydają się małe jak na tak wielkich roślinożerców. Dorosła samica nosorożca indyjskiego zajmuje średnio 2,9 km² ± 0,9 km². Samce zajmują nieznacznie większe obszary ze średnią 3,3 km². Nie wydaje się to zależeć od pory roku, bardziej już od jakości środowiska, w szczególności lubianych przez te zwierzęta traw. Nosorożec jawajski zajmuje niewielki obszary wobec ograniczonego zasięgu występowania Znacznie większy teren zdaje się zajmować nosorożec sumatrzański, 50 km². Nosorożec czarny zajmować potrafi aż 99 km² na nietrakcyjnych pod względem dostępności pokarmu równinach Serengeti. Zagęszczenia nosorożca białego wynoszą od 2,3 do 22,8 zwierzęcia na km², zależąc od pory roku i atrakcyjności siedliska, najwięcej na sawannach, nosorożca czarnego zaś między 0,01 a 1,5 na km²[60].
Dużą część doby tak wielkie zwierzęta muszą poświęcać na żerowanie, by dostarczyć wielkiemu ciału odpowiednią ilość pokarmu. Najwięcej czasu poświęca na żerowanie największy nosorożec biały, ponad połowę doby (50-60%). Prowadzi on też w największym stopniu z nosorożców dzienny tryb życia. Bardziej nocny jest nosorożec czarny. Nosorożec indyjski pozostaje pomiędzy nimi. Trudno wypowiadać się o nosorożcach jawajskim i sumatrzańskim, napotykanych rzadko i słabo poznanych. Ich obecnie w dużym stopniu nocna aktywność może wynikać z unikania kłusownictwa. Ponadto samice konsumują dłużej niż samce. Może to wynikać z dwóch powodów. Po pierwsze samice bywają w ciąży bądź karmią młode mlekiem, co przekłada się na zwiększone zapotrzebowanie na substancje odżywcze. Po drugie samce poświęcają czas na nieznane samicom aktywności, jak patrolowanie zajmowanego terenu w poszukiwaniu samic celem płodzenia dzieci i innych samców celem uniemożliwienia im tego samego, w efekcie czego dochodzi do kosztownych energetycznie walk. Aktywność zależy jednak od pory roku, zwłaszcza w przypadku nosorożca indyjskiego. W porze suchej, a więc od lutego do czerwca, nosorożce wykazują dwa szczyty żerowaniu w niskiej roślinności: przed świtem i około zmierzchu, z wyższej korzystają nieregularnie. W kwietniu, mimo upalnej pogody, jest dość sucho, więc łatwo pozbyć się nadmiaru ciepła z organizmu i zwierzęta nie spędzają aż tyle czasu w wodzie. Od czerwca do października nastaje wilgotna pora monsunowa, związana wobec dużej wilgotności i słabego wiatru z większym ryzykiem przegrzania się. Wtedy nosorożce więcej czasu taplają się w bajorkach. Trawy skubią po północy i późnym popołudniem do wieczora. Nieregularnie żerują na wyższej roślinności i odpoczywają. Gatunki afrykańskie odpoczywają najczęściej w gorące południe, spoczywając pod drzewem czy na szczycie wzgórza, wystawiając ciało na powiew wiatru, bądź też w płytkim zbiorniku wodnym[60].
Rozmieszczenie geograficzne
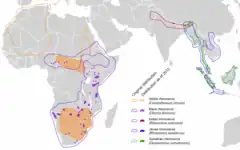
Rodzina obejmuje gatunki występujące w Afryce i Azji[64][130]. Niegdyś obejmowała szeroki połacie Eurazji i Amerykę Północną. Obecnie występuje jeszcze na południu, wschodzie i w centrum Afryki oraz na południu i południowym wschodzie Azji[60].
Ekologia

Nosorożcowate zamieszkują różnorodne siedliska[60].
Wielki nosorożec biały jest trawożercą. Zasiedla tereny trawiaste Afryki. Dzięki szerokim wargom i przystosowanym zębom potrafi nisko ścinać trawy, na wysokościach pozostawianych przez inne zwierzęta, jak bawół afrykański. Dzięki temu pobudza wzrost nowych traw, tworzy też strefy niskiej roślinności hamujące rozprzestrzenianie się pożarów. Preferencja w kierunku niskich traw może wiązać się z mniejszą ilością lignin, obficiej występujących w wyższych pędach. Do ulubionego pokarmu należą niskie Panicum coloratum, Urochloa mossabicensis, wyższe Panicum deustum i Panicum maximum rosnące pod drzewami, do których to w porze suchej dołącza Themeda triandra. Ścina trawy dość nisko, tworząc tzw. trawniki porównywane do pól golfowych, pobudzając wzrost niskiej trawy, bogatej w substancje odżywcze, stanowiące potem pokarm innych roślinożerców. Zwierzę unika natomiast lasów tropikalnych. Osobniki przenoszone na tereny górskie radziły sobie dobrze. Zwierzę potrafi nie pić kilka dni[60].
Nosorożec czarny odżywia się bardziej różnorodnie od większego krewniaka. W jego diecie naliczono ponad 200 gatunków roślin. Preferuje wyższą roślinność, zwłaszcza liście zrywane giętką górną wargą, nie pogardza też owocami. Na terenach suchych gustuje w sukulentach, bogatych w wodę. Szczególnie zjada wilczomlecz, nie bacząc na obecne w nim związki trujące dla innych zwierząt. Na wschodzie Afryki pożera zwłaszcza gatunek Euphorbia ingens, na zachodzie natomiast Euphorbia damariana. Pochłania rośliny wytwarzające lateks, który szkodzi człowiekowi nawet za dotknięciem. Tymczasem lateks widywano wokół ust nosorożca czarnego. Zjada Spirostachys africana. Trujące związki z jego diety ulegają rozkładowi w dużym jelicie ślepym zwierzęcia Łatwo wytrzymuje niedobór wody może się obyć bez niej kilka dni, czerpiąc wodę ze spożywanego pokarmu, jak to czynią inne pustynne ssaki, np. oryks. Dzięki temu zamieszkuje tereny pustynne, choć zazwyczaj pustyń unika. W Parku Narodowym Etosha 86,5% epizodów spotkania nosorożca czarnego miało miejsce w 16,5 powierzchni parku leżącego poniżej 10 km od wody, tylko zaś 1% dalej niż 15 km. Może nie pić 5 dni, aczkolwiek mając wybór idzie do wodopoju codziennie. Zaobserwowano, że utworzenie nowych wodopojów powiększyło jego zasięg na jednym z obszarów chronionych. Gatunek nie unika terenów górskich, osiągając wysokość 2900 m nad poziomem morza w Kenii[60].
Oba nosorożce afrykańskie słabo pływają. Nie potrafią przekroczyć większej rzeki i kiedy w wodzie stracą grunt pod nogami, mogą się utopić. Niekiedy rozgrzebują zagłębienia w ziemi, w których zmienia się woda, tworząc płytkie bajorka. w których potem zażywają kąpieli błotnych, nie korzystają jednak w ten sposób z głębszych zbiorników wodnych. Błoto pomaga pozbyć się pasożytów skórnych takich jak kleszcze, których pozostałości znajdywano w błocie opadłym z nosorożca. Zwierzęta niekiedy wychodzą ubłocone z bajorka, by potem ocierać się o drzewa, ścierając z siebie błoto wraz z ektopasożytami[60].
Zupełnie inne siedliska zamieszkują nosorożce azjatyckie. W odróżnieniu od swych afrykańskich krewnych świetnie pływają i dużą część czasu spędzają w wodzie. Zasiedlają równiny zalewowe rzek, zalewane okresowo lasy. Nosorożec indyjski zasiedla tereny monsunowe, nadrzeczne łąki porosłe wysokimi na 6-8 m trawami, także u podnóża Himalajów. Co roku ulewne deszcze pokrywają je metrową warstwą mułu, zasypującego mniej pożywne krzewy i przynoszącego minerały wspomagające wzrost żywiących nosorożce traw. Świetnie pływając, nosorożec żywi się także roślinami wodnymi. Zasiedla tereny bogate w Saccharum spontaneum, nie tylko łatwostrawne, ale i pobudzane do wzrostu przez zgryzanie czy zalewanie. Przez większą część roku gatunek ten stanowi ponad połowę pokarmu ssaka. Mniej strawne są za to Narenga porphyracoma i Themeda arundinacea, co skutkuje mniejszymi zagęszczeniami nosorożców indyjskich w zdominowanych przez nie okolicach. Wysokich na 6-8 „słoniowych traw” unika, jedząc zazwyczaj wyłącznie młode odrosty na przykład po pożarze. Niemniej ekstremalne powodzie nie służą nosorożcom, które potrzebują jednak jakiegoś lądu. W razie znacznych powodzi przenoszą się do niezalewanych lasów. Większość rezerwatów zdominowanych jest jednak przez unikaną przez nosorożce indyjskie z powodu tanin Shorea robusta. W lasach tych bogato rosną również inne nielubiane przez nosorożce rośliny, mało jest też wody. Nosorożec indyjski rzadko oddala się od wody na dalej niż 2 km. W Nepalu odnotowano 183 gatunki roślin, którymi się żywi, należące do ponad pół setki rodzin, wśród nich turzyce, paprotniki, rośliny wodne, pnącza, zioła. Na inne prócz traw gatunki roślin zwierzę przerzuca się w porze suchej, zwłaszcza Litsea monopetala i Mallotus philippinensis. Wtedy to trawy robią się bardziej suche. Obserwowano nosorożce gustujące w papryczkach chili, ale niekoniecznie już w gorczycy, konsumowanej przez nie rzadko[60].
Nosorożec jawajski prawdopodobnie zasiedlał także lasy mangrowe i zalewowe, zanim wyginał w Sundarbanach. Nosorożec sumatrzański preferował prawdopodobnie podobne, ale wyżej położone lasy. Niemniej obecny zasięg występowania obu gatunków jest ograniczony i może obejmować raczej obszary mniej zagrożone kłusownictwem niż siedliska rzeczywiście preferowane przez nosorożce. Pierwotny tropikalny las nie wydaje się najlepszym siedliskiem dla nosorożca jawajskiego. Żeruje on głownie w lesie wtórnym, na polanach, miejscach po upadłych drzewach, gdzie następuje intensywny wzrost nowych roślin, produkujących dużo białka, a niewiele toksycznych wtórnych metabolitów. Nosorożce jawajskie prawdopodobnie skorzystały na zniszczeniu lasu po erupcji Krakatau pod koniec XIX wieku, po której zaczął odrastać nowy las. Obecnie zdaje się on już zbyt dojrzały i liczebność nosorożców maleje. Dominująca w podszycie Arecia pinnata nie stanowi dla nich dogodnego pokarmu, w przeciwieństwem do Acacia pennata, Bambusia, Calamus, Cyathea, Strychnos nux-vomica. Ostatni z wymienionych jest trujący, aczkolwiek nie wiadomo, czy nosorożce zjadają tylko mięsiste owoce, czy także inne, toksyczne, części tych roślin[60].
Dużą część czasu, nawet 8 godzin na dobę, w porze deszczowej, ciepłej i wilgotnej, kiedy to termoregulacja staje się trudna, a ryzyko przegrzania organizmu rośnie, nosorożce spędzają w wodnych bajorkach. Potrafi gromadzić się w nich nawet ponad 30 sztuk nosorożca indyjskiego, który nie wykazuje w nich terytorializmu. Taplanie się spełnia zarazem funkcję społeczną, nie tylko wspólne, albowiem nosorożec nie krępuje się oddawać moczu do wody, w której się kąpie, pozostawiając w niej swój zapach. Nosorożec sumatrzański, kiedy brakuje niewielkich bajorek, korzysta z lasów mangrowych i większych zbiorników, ale zwykle pozostaje przy brzegu, gdzie jest więcej błota, i drąży w nim, zmieniając brzegi zbiornika. W chłodniejszej porze roku czas spędzany w wodzie jest znacznie krótszy[60].
Nosorożce azjatyckie zasiedlają tereny znacznie liczniej zamieszkiwane przez człowieka niż afrykańskie sawanny nosorożców czarnych czy białych. W efekcie pojawiają się też w środowiskach zmienionych ręką człowieka, w tym wśród upraw pszenicy, kukurydzy, ryżu czy soczewicy. Uznaje się je za szkodniki. Nosorożce jawajskie zapuszczają się także na plantacje herbaty, gdzie ludzie, nie bacząc na ich status gatunku krytycznie zagrożonego wyginięciem, zabijają je[60].
Wszystkie gatunki azjatyckie korzystają z lizawek solnych. Nosorożec sumatrzański używa ich raz w tygodniu. Nosorożec indyjski znajduje je w lasach Shorea robusta[60].
Zagrożenia i ochrona

Nosorożcowate niegdyś zamieszkiwały szerokie połacie wielu kontynentów. Obecnie przetrwało 5 gatunków, z których wszystkim zagraża wyginięcie, choć w różnym stopniu[60].
W Afryce gęstość zaludnienia na wielu zamieszkiwanych przez nosorożce obszarach jest cały czas niewielka, a działalność człowieka ograniczona. Obszary zachowanej dzikiej przyrody osiągają niekiedy wielkość całych europejskich krajów. Głównym zagrożeniem jest raczej kłusownictwo celem pozyskania cennych rogów, sprzedawanych po 30000 dolarów za kilogram. Kłusownictwu sprzyja niepewna sytuacja polityczna w wielu afrykańskich krajach, miejscowe przewroty i powstania, a także związana z nimi znaczna dostępność broni. Kraje południa Afryki podjęły pod koniec XX wieku dość kontrowersyjną akcję odcinania nosorożcom rogów w nadziei, że zniechęci to kłusowników. Akcja ta nie odniosła istotnego sukcesu. Z jednej strony pozbawienie rogu pojedynczego zwierzęcia wiązała się z kosztem półtora tysięcy dolarów, co pozwalało na ochronę w ten sposób jedynie niewielkiej populacji. Zwolennicy akcji podkreślali, że wystarczy obciąć rogi tylko części populacji, a już wtedy kłusownictwo stanie się nieopłacalne, jednakże kłusownicy i tak zabijali nosorożce, niekiedy nie sprawdzając przed zabiciem, czy zwierzę w ogóle ma róg. Poza tym rogi, zbudowane z keratyny jak włosy czy paznokcie, dość szybko odrastają, a wysoka cena rynkowa czyniła opłacalnym pozyskanie już młodego, jednorocznego odrostu. Ponadto zaczęły pojawiać się dane o zwiększonej śmiertelności młodych bezrogich samic z powodu hienowatych i lwów. W efekcie z tej praktyki wycofano się[60].
Nosorożec czarny zasiedlał w przeszłości dużą część Afryki, licząc przynajmniej 300000, a być może nawet milion osobników. Później jego zasięg występowania zmniejszył się gwałtownie, w dużej mierze na skutek polować przez kłusowników. W pewnym momencie pozostały już tylko niewielki populacje w kilku objętych ochroną miejscach. Jednakże zwiększyły one następnie swą liczebność, osiągając maksymalny rozmiar w danym miejscu, w efekcie czego nosorożce zaczęto przenosić w inne miejsca. Szczególnie udała się ochrona w Parku Narodowym Etosha w Namibii, gdzie niewielka grupka rozrosła się w 800 sztuk[60].
Mniej szczęścia miał północny podgatunek nosorożca białego. Do końca XX wieku pozostały niewielkie populacje, a niedługo później ostatnie żyjące na wolności osobniki zdechły. Wobec niewielkiej liczby zwierząt pozostałych jeszcze w ogrodach zoologicznych nie ma szans na przywrócenie podgatunku. O skraj zagłady otarł się także podgatunek południowy tego samego gatunku, w najgorszym okresie licząc jakiś 100 zwierząt w jednym z rezerwatów na południu kontynentu około 1900 roku. Liczba osobników rosła jednak w rezerwacie Hluhluwe-Umfolozi, z którego przenoszono je w inne miejsca. W efekcie liczebność przekroczyła 20000 sztuk, a status gatunku spadł do NT (Near Threatened)[60].
Zupełnie inaczej wygląda sytuacja w państwach azjatyckich, cechujących się dużym zagęszczeniem populacji człowieka rozumnego i ograniczonymi jedynie niezniszczonymi przez człowieka siedliskami. W sumie wszystkie populacja liczą niecałe 2700 osobników, przy czym jedynie dwie populacje liczą ponad setkę nosorożców. Z drugiej strony miejscowa ludność zazwyczaj nie ma dostępu do broni. Pojedyncze przypadki, kiedy wybuchło lokalne powstanie i w rękach miejscowej ludności pojawiła się broń, wiązały się z wytrzebieniem lokalnych populacji nosorożców[60].
Nosorożec indyjski, niegdyś liczący prawie pół miliona sztuk, a potem uważany za zwierzę pozostające pod ochroną królewską, ostał się jedynie w kilku miejscach, w tym w parkach narodowych Kaziranga i Royal Chitwan. Był z nich potem przenoszony w inne miejsca. W niektórych w ochronie uczestniczy miejscowa ludność, czerpiąca zyski z turystyki[60].
Znacznie gorzej wygląda sytuacja nosorożców sumatrzańskiego i jawajskiego. Nosorożec sumatrzański ostał się w kilku miejscach, w tym na północy Borneo, jednak żadna populacja nie przekracza 50 zwierząt. Spektakularną porażką zakończyła się próba hodowli i rozrodu w niewoli, rozpoczęta odłowem populacji skazanej w naturze na wymarcie. W niewoli ssaki te nie tylko nie chciały rozmnażać się, ale dość szybko umierały. Kilka młodych narodziło się co prawda w zoo w Cincinatti, ale nie osiągnięto rozrodczości obserwowanej w naturze. Jednym sposobem utrzymania istnienia gatunku są próby ochrony go ww jego naturalnym środowisku we współpracy z miejscową ludnością[60].
Jeszcze gorzej wygląda sytuacja nosorożca jawajskiego, uznawanego za najbardziej zagrożonego ze ssaków. Pozostały wyłącznie dwie populacje. Mniejsza z nich żyła na granicy Wietnamu z Kambodżą. Jednakże liczebność jej szacowano na zaledwie kilka zwierząt, a ostatni raz napotkano osobnika martwego, zabitego przez kłusowników. W efekcie uznano, że populacja wymarła. Natomiast w Parku Narodowym Ujung Kulon na Jawie żyje około 50 sztuk. Tak mała liczebność w jednym jedynym miejscu rodzi istotne problemy, narażając gatunek na wymarcie z powodu chorób czy wybuchu wulkanu[60].
W kulturze
Nosorożcowate odgrywają istotną rolę w kulturze. Europejczycy zapoznali się z istnieniem takiego zwierzęcia w 1515, kiedy to Albrecht Durer przedstawił je na drzeworycie, aczkolwiek w sposób odbiegający od rzeczywistych cech zwierząt. Wcześniej już w europejskiej kulturze obecny był mityczny jednorożec, aczkolwiek stworzenie znaczniej bardziej zwinne od przygrubego nosorożca. Nosorożce bywały też uznawane za święte zwierzęta w Azji. W Nepalu były własnością władcy, a zabiciem nosorożca państwo zajmowało się pieczołowiciej niż zabójstwem człowieka[60].
W Azji nosorożce wykorzystuje się w medycynie tradycyjnej, zwłaszcza chińskiej, dokładniej rogi, pomimo braku dowodów na działanie. Wbrew rozpowszechnionemu poglądowi róg nosorożca nie jest wykorzystywany jako afrodyzjak. Azjaci wykorzystują go raczej do leczenia gorączki. Stosują przy tym bardzo niewielkie ilości sproszkowanego rogu, tak że zapotrzebowanie jest niewielkie, a pojedynczy róg mógłby zaopatrzyć cały kraj na okres roku. Pojawiają się również informacje, jakoby róg miał leczyć raka. Pomimo tego cena na czarnym rynku może wynosić 30000 za kg i nosorożce padają ofiarami kłusowników. Cena rogów gatunków azjatyckich przekracza tutaj ceny rogów nosorożców z Afryki, być może na skutek mniejszych rozmiarów rogów[60].
Uwagi
- ↑ Definicja taksonu nie opiera się na nazwie rodzajowej.
- 1 2 3 4 5 6 Definiacja taksonu nie opiera się na nazwie rodzajowej.
- ↑ Typ nomenklatoryczny: †Aceratherium Kaup, 1832.
- ↑ Typ nomenklatoryczny: †Atelodus Pomel, 1853 (= †Coelodonta Bronn, 1831).
- ↑ Typ nomenklatoryczny: †Teleoceras Hatcher, 1894.
- ↑ Niepoprawna późniejsza pisownia Rhinoceroscidæ Burnett, 1829.
- ↑ Typ nomenklatoryczny: Dicerorhinus Gloger, 1841.
- ↑ Typ nomenklatoryczny: Diceros J.E. Gray, 1821.
- ↑ Typ nomenklatoryczny: Ceratotherium J.E. Gray, 1868.
- ↑ Typ nomenklatoryczny: †Ronzotherium Aymard, 1854.
- ↑ Typ nomenklatoryczny: †Turkanatherium Deraniyagala, 1951 (= †Aceratherium Kaup, 1832).
- ↑ Typ nomenklatoryczny: †Lartetotherium Ginsburg, 1974.
- ↑ Typ nomenklatoryczny: †Trigonias F.A. Lucas, 1900.
- ↑ Typ nomenklatoryczny: Punjabitherium Khan, 1971 (= Rhinoceros Linnaeus, 1758.)
- ↑ Typ nomenklatoryczny: †Coelodonta Bronn, 1831.
- ↑ Typ nomenklatoryczny: †Alicornops Ginsburg & Guérin, 1979.
Przypisy
- 1 2 J.E. Gray. On the Natural Arrangment of Vertebrose Animals. „The London Medical Repository”. 15, s. 306, 1821. (ang.).
- ↑ J.K.W. Illiger: Prodromus systematis mammalium et avium: additis terminis zoographicis utriusque classis, eorumque versione germanica. Berolini: Sumptibus C. Salfeld, 1811, s. 97. (łac.).
- ↑ C.S. Rafinesque: Analyse de la nature, or, Tableau de l’univers et des corps organisés. Palerme: Aux dépens de l’auteur, 1815, s. 56. (fr.).
- ↑ J.E. Gray. An Outline of an Attempt at the Disposition of Mammalia into Tribes and Families, with a List of the Genera apparently appertaining to each Tribe. „Annals of Philosophy”. New Series. 10, s. 337–344, 1825. (ang.).
- ↑ J. Brookes: A catalogue of the Anatomical and Zoological Museum of Joshua Brookes. Cz. 16. London: Richard Taylor, 1828, s. 74. (ang.).
- ↑ G.T. Burnett. Illustrations of the Quadrupeda, or Quadrupeds, being the arrangement of the true four-footed Beasts indicated in outline. „Quarterly Journal of Science, Literature and the Arts”. 28, s. 352, 1829. (ang.).
- ↑ Ch.L. Bonaparte. Synopsis Vertebratorum Systematis. „Nuovi annali delle scienze naturali”. 2, s. 113, 1838. (łac.).
- ↑ R.-P. Lesson: Nouveau Tableau du Règne Animal: Mammifères. Paris: Arthus Bertrand, 1842, s. 158. (fr.).
- ↑ R. Owen: Odontography: or a treatise on the comparative anatomy of the teeth, their physiological relations, mode of development and microscopic structure, in the vertebrate animals. London: Hippolyte Baillier, 1845, s. 587. (ang.).
- ↑ J.-Ch. Chenu & A.G. Desmarest: Pachydermes. W: J.-Ch. Chenu: Encyclopédie d’histoire naturelle; ou, traité complet de cette science d’après les travaux des naturalistes les plus éminents de tous les pays et de toutes les époques: Buffon, Daubenton, Lacépède, G. Cuvier, F. Cuvier, Geoffroy Saint-Hilaire, Latreille, De Jussieu, Brongniart, etc. Cz. 7: Pachydermes, ruminants, édentés, cétacés, marsupiaux et monotrèmes. Paris: Maresq, 1857, s. 1. (fr.).
- ↑ E. Haeckel: Generelle morphologie der organismen. Allgemeine grundzüge der organischen formen-wissenschaft, mechanisch begründet durch die von Charles Darwin reformirte descendenztheorie. Cz. 2. Berlin: G. Reimer, 1866, s. clviii. (niem.).
- ↑ J.E. Gray. Observations on the Preserved Specimens and Skeletons of the Rhinocerotidæ in the Collection of the British Museum and Royal College of Surgeons, including the Descriptions of Three New Species. „Proceedings of the Zoological Society of London”. 1867, s. 1005, 1867. (ang.).
- ↑ T.N. Gill. Arrangement of the Families of Mammals; with Analytical Tables. „Smithsonian Miscellaneous collections”. 11, s. 12, 1872. (ang.).
- ↑ J.E. Gray: Hand-list of the edentate, thick-skinned and ruminant mammals in the British Museum. London: The Trustees, 1873, s. 45. (ang.).
- ↑ E. Blyth. Catalogue of mammals and birds of Burma. „The Journal of the Asiatic Society of Bengal”. Supplement. 44 (2), s. 50, 1875. (ang.).
- 1 2 von Brandt 1878 ↓, s. 25.
- ↑ von Brandt 1878 ↓, s. 33.
- ↑ von Brandt 1878 ↓, s. 41.
- ↑ von Brandt 1878 ↓, s. 51.
- 1 2 3 4 L. Dollo. Rhinocéros vivants et fossiles. „Revue des questions scientifiques”. 17, s. 295, 1885. (fr.).
- ↑ F. Ameghino. Contribución al conocimiento de los mamiferos fósiles de la República Argentina. „Acta de la Academia Nacional de Ciencias Exactas”. 6, s. 894, 1889. (hiszp.).
- ↑ K.A. von Zittel: Traité de paléontologie. T. 4: Vertebrata (Mammalia). Munchen and Leipzig: R. Oldenburg, 1891, s. 288. (niem.).
- ↑ H.F. Osborn. The Extinct Rhinoceroses. „Memoirs of the American Museum of Natural History”. 1 (3), s. 121, 1898. (ang.).
- ↑ Osborn 1900 ↓, s. 249.
- ↑ Osborn 1900 ↓, s. 262.
- ↑ O.P. Hay. Bibliography and catalogue of the fossil vertebrata of North America. „Bulletin of the United States Geological Survey”. 179, s. 646, 1902. (ang.).
- ↑ T.S. Palmer. Index Generum Mammalium: a List of the Genera and Families of Mammals. „North American Fauna”. 23, s. 768, 1904. (ang.).
- ↑ H. Winge: Jordfundne og nulevende hovdyr (Ungulata) fra Lagoa Santa, Minas Geraes, Brasilien: med udsigt over hovdyrenes indbyrdes slægtskab. Kjøbenhavn: H. Hagerup, 1906, s. 71, seria: E Museo Lundii. Triede Bind. Første halvbind. (duń.).
- ↑ W.K. Gregory & J.K. Mosenthal: Outline classification of the Mammalia recent and extinct. W: H.F. Osborn: The Age of Mammals in Europe, Asia and North America. New York: The Macmillan Company, 1910, s. 557. (ang.).
- ↑ S. Breuning. Beiträge zur Stammesgeschichte der Rhinocerotidae. „Verhandlungen der Zoologische und Botanische Gesellschaft Wien”. 73, s. 120, 1923. (niem.).
- ↑ Ringström 1924 ↓, s. 5.
- ↑ Ringström 1924 ↓, s. 97.
- ↑ V.T. Zammarano: Le colonie italiane di diretto dominio. Fauna e caccia. A cura di. Rome: Ministero delle colonie, Ufficio studi e propaganda, 1930, s. 165. (wł.).
- ↑ W.O. Dietrich. Zur Entwicklungsmechanik des Gebisses der afrikanischen Nashörner. „Centralblatt für Mineralogie, Geologie und Paläontologie”. 1942, s. 297–300, 1942. (niem.).
- ↑ M. Kretzoi. Alttertiäre Perissodactylen aus Ungarn. „Annales historico-naturales Musei Nationalis Hungarici”. 33, s. 87–98, 1940. (niem.).
- ↑ G.G. Simpson. The principles of classification and a classification of mammals. „Bulletin of the American Museum of Natural History”. 85, s. 142, 1945. (ang.).
- ↑ M. Friant. Recherches sur le télencéphale des Rhinoceridae. „Proceedings of the Zoological Society of London”. 1950 (2), s. 337, 1950. DOI: 10.1111/j.1096-3642.1950.tb00953.x. (ang.).
- 1 2 P.E.P. Deraniyagala. A hornless rhinoceros from the Mio-Pliocene deposits of East Africa. „Spolia Zeylanica”. 26, s. 24, 1951. (ang.).
- ↑ J.R. Ellerman, T.C.S. Morrison-Scott & R.W. Hayman: Southern African mammals 1758 to 1951: a reclassification. London: British Museum (Natural History), 1953, s. 162. (ang.).
- ↑ P.E.P. Deraniyagala. The Extinct and Living Subfamilies of African Rhinocerotidae. „Spolia Zeylanica”. 27, s. 13, 1953. (ang.).
- ↑ Heißig 1972 ↓, s. 16.
- ↑ Heißig 1972 ↓, s. 58.
- ↑ Heißig 1972 ↓, s. 59.
- ↑ Heißig 1972 ↓, s. 71.
- 1 2 H. Loose. Pleistocene Rhinocerotidae of W. Europe with reference to the recent two-horned species of Africa and S.E. Asia. „Scripta Geologica”. 33, s. 7, 1975. (ang.).
- 1 2 C.P. Groves. Phylogeny of the living species of rhinoceros. „Zeitschrift für Zoologische Systematik und Evolutionsforschung”. 21 (4), s. 312, 1983. DOI: 10.1111/j.1439-0469.1983.tb00297.x. (ang.).
- ↑ K. Heißig: Rhinocerotidae. W: D.R. Prothero & R.M. Schoch (redaktorzy): The evolution of perissodactyls. New York: Oxford University Press, 1989, s. 405, 406, seria: Oxford Monographs on Geology and Geophysics, 15. ISBN 978-0195060393. (ang.).
- ↑ Prothero i Schoch 1989 ↓, s. 534.
- 1 2 3 Prothero i Schoch 1989 ↓, s. 536.
- ↑ Guérin 1989 ↓, s. 5.
- 1 2 Guérin 1989 ↓, s. 10.
- ↑ Kalandadze i Rautian 1992 ↓, s. 115.
- 1 2 3 Kalandadze i Rautian 1992 ↓, s. 116.
- ↑ E. Cerdeño. Cladistic analysis of the family Rhinocerotidae (Perissodactyla). „American Museum Novitates”. 3143, s. 23, 1995. (ang.).
- ↑ McKenna i Bell 1997 ↓, s. 482.
- ↑ McKenna i Bell 1997 ↓, s. 483.
- ↑ McKenna i Bell 1997 ↓, s. 484.
- ↑ P.-O. Antoine. Phylogénie et évolution des Elasmotheriina (Mammalia, Rhinocerotidae). „Mémoires du Muséum national d’Histoire naturelle”. 188, s. 37, 2002. (fr.).
- 1 2 Nazwy zwyczajowe za: W. Cichocki, A. Ważna, J. Cichocki, E. Rajska-Jurgiel, A. Jasiński & W. Bogdanowicz: Polskie nazewnictwo ssaków świata. Warszawa: Muzeum i Instytut Zoologii PAN, 2015, s. 166–167. ISBN 978-83-88147-15-9. (pol. • ang.).
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 E. Dinerstein, Family Rhinocerotidae (Rhinoceroses), [w:] D.E. Wilson & R.A. Mittermeier (redaktorzy), Handbook of the Mammals of the World, t. 2. Hoofed Mammals, Barcelona: Lynx Edicions, 2011, s. 144-, ISBN 978-84-96553-77-4 (ang.).
- ↑ Gruboskórne zwierzęta. W: Encyklopedyja powszechna. T. 10: Glin.–Guis.. Warszawa: Samuel Orgelbrand, 1862, s. 793-794.
- ↑ N. Upham, C. Burgin, J. Widness, M. Becker, C. Parker, S. Liphardt, I. Rochon & D. Huckaby: Treeview of Mammalian Taxonomy Hierarchy. [w:] ASM Mammal Diversity Database (Version 1.11) [on-line]. American Society of Mammalogists. [dostęp 2023-12-10]. (ang.).
- ↑ Class Mammalia. W: Lynx Nature Books: All the Mammals of the World. Barcelona: Lynx Edicions, 2023, s. 637–638. ISBN 978-84-16728-66-4. (ang.).
- 1 2 C.J. Burgin, D.E. Wilson, R.A. Mittermeier, A.B. Rylands, T.E. Lacher & W. Sechrest: Illustrated Checklist of the Mammals of the World. Cz. 2: Eulipotyphla to Carnivora. Barcelona: Lynx Edicions, 2020, s. 390. ISBN 978-84-16728-35-0. (ang.).
- ↑ J.S. Zijlstra, Elasmotheriinae, Hesperomys project (Version 23.8.1), DOI: 10.5281/zenodo.7654755 [dostęp 2023-12-10] (ang.).
- ↑ J.J. Kaup. Ueber Rhinoceros incisivus Cuv., and eine neue Art, Rhinoceros Schleiermacheri. „Isis von Oken”. Jahrgang 1832, s. 904, 1832. (niem.).
- 1 2 M. Kretzoi. Bemerkungen zum System der Nachmiozänen Nashorn-Gattungen. „Földtani közlöny”. 72 (4–12), s. 311, 1942. (niem.).
- ↑ L. Ginsburg & C. Guérin. Sur l’origine et l’extension stratigraphique du petit Rhinocerotide miocene Acerotherium (Alicornops) simorrense (Lartet 1851) nov. subgen. „Comptes Rendus Sommaires de la Société Géologique de France”. 3, s. 114, 1979. (fr.).
- ↑ H.E. Wood, 2nd. Some early Tertiary rhinoceroses and hyracodonts. „Bulletins of American paleontology”. 13 (50), s. 232, 1927. (ang.).
- ↑ E.D. Cope. On some new extinct Mammalia from the Tertiary of the Plains. „Paleontological bulletin”. 14, s. 1, 1873. (ang.).
- ↑ C. Forster Cooper. New genera and species of mammals from the Miocene deposits of Baluchistan.—Preliminary notice. „The Annals and Magazine of Natural History”. Eight series. 16 (95), s. 408, 1915. (ang.).
- ↑ O. Roger. Wirbeltierreste aus dem Obermiocän der bayerisch-schwäbisclien Hochebene. „Bericht des Naturwissenschaftlichen Vereins für Schwaben und Neuburg (a.V.) in Augsburg”. Veröifentlieht im Jahre 1904, s. 13, 1904. (niem.).
- ↑ P. Antoine & J. Welcomme. A new rhinoceros from the Lower Miocene of the Bugti Hills, Baluchistan, Pakistan: The earliest elasmotheriine. „Palaeontology”. 43 (5), s. 798, 2000. DOI: 10.1111/1475-4983.00150. (ang.).
- ↑ K. Heißig. Paläontologische und geologische Untersuchungen im Tertiär von Pakistan 5. Rhinocerotidae (Mamm.) aus den unteren und mittleren Siwalik-Schichten. „Abhandlungen der Bayerischen Akademie der Wissenschaften (mathematisch-naturwissenschaftliche Klasse)”. Neue Folge. 152, s. 50, 1972. (niem.).
- ↑ D.A. Hooijer. A new rhinoceros from the late Miocene of Loperot, Turkana district, Kenya. „Bulletin of the Museum of Comparative Zoology at Harvard College”. 142 (3), s. 342, 1971. (ang.).
- ↑ Ringström 1924 ↓, s. 26.
- ↑ H.G. Bronn. Ueber die fossilen Zähne eines neuen Geschlechtes aus der Dickhäuter-Ordnung, Coelodonta, Höhlenzahn. „Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde”. 2, s. 60, 1831. (niem.).
- ↑ W.O. Dietrich. Neue Nashornreste aus Schwaben (Diaceratherium tomerdingensis n. g. n. sp.). „Zeitschrift für Säugetierkunde”. 6, s. 203, 1931. (niem.).
- ↑ O.Ch. Marsh. Notice of new Tertiary mammals, IV. „The American journal of science and arts”. Third series. 9 (51), s. 242, 1875. (ang.).
- ↑ von Brandt 1878 ↓, s. 48.
- ↑ M. Crusafont i Pairó, J.F. De Villalta & J.T. Santonja. El burdigaliense continental de la cuenca del Vallés-Penedés. „Memorias y comunicaciones del Instituto Geológico, Diputación Provincial de Barcelona”, s. 152–156, 1955. (ang.).
- ↑ J.G. Fischer von Waldheim. La notice d’un animal fossile de Sibirie inconnu aux naturalistes. „Programme d’invitation a la seance publique de la Societe Imperiale des naturalistes de Moscou”. 4, s. 23, 1808. (fr.).
- ↑ D. Geraads & S. Zouhri. A new late Miocene elasmotheriine rhinoceros from Morocco. „Acta Palaeontologica Polonica”. 66 (4), s. 754, 2021. DOI: 10.4202/app.00904.2021. (ang.).
- ↑ Abel 1910 ↓, s. 20.
- ↑ H.E. Wood, 2nd. Rhinoceroses from the Thomas Farm Miocene of Florida. „Bulletin of the Museum of Comparative Zoology at Harvard College”. 130 (5), s. 364, 1964. (ang.).
- ↑ E.H. Colbert. A new rhinoceros from the Siwalik beds of India. „American Museum novitates”. 749, s. 1, 1934. (ang.).
- ↑ Prothero 2005 ↓, s. 70.
- ↑ L.B. Albright. Ungulates of the Toledo Bend local fauna (late Arikareean, early Miocene), Texas Coastal Plain. „Bulletin of the Florida Museum of Natural History”. Biological Sciences. 42 (1), s. 22, 1999. (ang.).
- ↑ M. Crusafont i Pairó & J.F. de Villalta Comella. Sobre un interesante rinoceronte (Hispanotherium nov. gen.) del Mioceno del valle de Manzanares. Nota preliminar. „Las Ciencias”. 12 (4), s. 867, 1947. (hiszp.).
- ↑ L. Ginsburg & K. Heißig: Hoploaceratherium, a new generic name for ‘Aceratherium’ tetrcidactylum. W: D.R. Prothero & R.M. Schoch (redaktorzy): The evolution of perissodactyls. New York: Oxford University Press, 1989, s. 418, seria: Oxford Monographs on Geology and Geophysics, 15. ISBN 978-0195060393. (ang.).
- ↑ Huang i Yan 1983 ↓, s. 226.
- ↑ Ringström 1924 ↓, s. 147.
- ↑ Е.И. Беляева: Семейство Rhinocerotidae. W: Э.А. Вангенгейм, Е.И. Беляева, B.E. Гарутт, Е.Л. Дмитриева & В.C. Зажигин: Млекопитающие эоплейстоцена западного Забайкалья. Москва: Издательство «Наука», 1966, s. 133. (ros.).
- ↑ E. Aguirre & C. Guérin. Première découverte d’un Iranotheriinae (Mammalia, Perissodactyla, Rhinocerotidae) en Afrique: Kenyatherium bishopi nov. gen. nov. sp. de la formation vallésienne (Miocène supérieur) de Nakali (Kenya). „Estudios Geológicos”. 30 (3), s. 232, 1974. (fr.).
- ↑ L. Ginsburg. Les rhinocerotides du Miocene de Sansan (Gers). „Comptes rendus hebdomadaires de l’Académie des Sciences”. Série D. 278 (5), s. 597, 1974. (fr.).
- ↑ E.L. Troxell. A study of Diceratherium and the diceratheres. „The American journal of science”. Fifth series. 2 (10), s. 206, 1921. (ang.).
- ↑ K. Heißig. Die Rhinocerotidae (Mammalia) aus der oberoligozänen Spaltenfüllung von Gaimersheim bei Ingolstadt in Bayern und ihre phylogenetische Stellung. „Abhandlungen der Bayerischen Akademie der Wissenschaften (mathematisch-naturwissenschaftliche Klasse)”. Neue folge. 138, s. 90, 1969. (niem.).
- ↑ I.X. Giaourtsakis: The fossil record of rhinocerotids (Mammalia: Perissodactyla: Rhinocerotidae) in Greece. W: E. Vlachos (red.): Fossil Vertebrates of Greece. Cz. 2: Laurasiatherians, Artiodactyles, Perissodactyles, Carnivorans, and Island Endemics. Cham: Springer, 2022, s. 441. ISBN 978-3-030-68441-9. (ang.).
- ↑ D. Becker, P.-O. Antoine & O. Maridet. A new genus of Rhinocerotidae (Mammalia, Perissodactyla) from the Oligocene of Europe. „Journal of Systematic Palaeontology”. 11 (8), s. 950, 2013. DOI: 10.1080/14772019.2012.699007. (ang.).
- ↑ P.-O. Antoine, M.C. Reyes, N. Amano, A.P. Bautista, Ch.-H. Chang, J. Claude, J. de Vos & T. Ingicco. A new rhinoceros clade from the Pleistocene of Asia sheds light on mammal dispersals to the Philippines. „Zoological Journal of the Linnean Society”. 194 (2), 2022. DOI: 10.1093/zoolinnean/zlab009. (ang.).
- ↑ 陈冠芳 / G. Chen. 宁夏中宁一板齿犀化石 / A new genus of Iranotheriinae of Ningxia. „古脊椎动物学报 / Vertebrata PalAsiatica”. 15 (2), s. 143, 1977. (chiń. • ang.).
- ↑ C. Guérin & M. Pickford. Ougandatherium napakense nov. gen. nov. sp., le plus ancien Rhinocerotidae Iranotheriinae d’Afrique. „Annales de Paléontologie”. 89 (1), s. 31, 2003. DOI: 10.1016/S0753-3969(03)00004-1. (fr.).
- ↑ D.A. Hooijer. A rhinoceros from the Late Miocene of Fort Ternan, Kenya. „Zoologische Mededelingen”. 43 (6), s. 78, 1968. (ang.).
- ↑ L.G. Tanner & L.D. Martin. New Rhinocerotids from the Oligocene of Nebraska. „Life Science Miscellaneous Publications of the Royal Ontario Museum, Toronto”, s. 212, 1976. (ang.).
- ↑ E.D. Cope. A New Genus of Rhinocerontidæ. „The American naturalist”. 14, s. 540, 1880. (ang.).
- ↑ H. Killgus. Ünterpliozäne Sauger aus China. „Paläontologische Zeitschrift”. 5 (3), s. 251, 1923. DOI: 10.1007/BF03160375. (ang.).
- ↑ L. Pandolfi. Persiatherium rodleri, gen. et sp. nov. (Mammalia, Rhinocerotidae) from the upper Miocene of Maragheh (northwestern Iran). „Journal of Vertebrate Paleontology”. 36 (1), s. 2, 2016. DOI: 10.1080/02724634.2015.1040118. (ang.).
- ↑ Z. Yang. On a Miocene mammalian fauna from Shantung. „Bulletin of the Geological Society of China”. 17 (2), s. 214, 1937. DOI: 10.1111/j.1755-6724.1937.mp17002007.x. (ang.).
- ↑ O. Roger. Wirbelthierreste aus dem Dinotheriensande der bayerisch-schwäbischen Hochebene. „Bericht des Naturwissenschaftlichen Vereins für Schwaben und Neuburg (a.V.) in Augsburg”. Veröffentlicht im Jahre 1898, s. 25, 1898. (niem.).
- ↑ W.D. Matthew. Critical observations on the phylogeny of the rhinoceroses. „University of California Publications, Bulletin of the Department of Geological Sciences”. 20 (1), s. 7, 1931. (ang.).
- ↑ A. Koch. Prohyracodon orientalis, ein neues Ursaugethier aus den Mitteleocänen Schichten Siebenbürgens. „Természetrajzi Füzetek kiadja a Magyar nemzeti Muzeum”. 20, s. 481, 1897. (niem.).
- ↑ K. Heißig. Prosantorhinus pro Brachypodella Heissig 1972 (Rhinocerotidae, Mammalia) (= non Brachypodella Beck 1837 <Gastropoda>). „Mitteilungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie”. 14, s. 37, 1974. (niem.).
- ↑ Abel 1910 ↓, s. 10.
- ↑ A. Aymard. Des terrains fossilifères du bassin supérieur de la Loire. „Comptes rendus hebdomadaires de l’Académie des Sciences”. 38, s. 675, 1854. (fr.).
- ↑ D. Geraads: Rhinocerotidae. W: L. Werdelin & W.J. Sanders (redaktorzy): Cenozoic Mammals of Africa. Berkeley: University of California Press, 2010, s. 682. ISBN 978-0-520-25721-4. (ang.).
- ↑ N. Handa, M. Nakatsukasa, Y. Kunimatsu & H. Nakaya. A new Elasmotheriini (Perissodactyla, Rhinocerotidae) from the upper Miocene of Samburu Hills and Nakali, northern Kenya. „Geobios”. 50 (3), s. 199, 2017. DOI: 10.1016/j.geobios.2017.04.002. (ang.).
- ↑ Huang i Yan 1983 ↓, s. 223.
- ↑ T.J. Ringström. Sinotherium lagrelii, a new fossil rhinocerotid from Shansi. „Bulletin of the Geological Survey of China”. 5 (1), s. 91, 1923. (ang.).
- ↑ Prothero 2005 ↓, s. 58.
- ↑ M. Kretzoi. Präokkupierte und durch ältere zu ersetzende Säugetiernamen. „Földtani közlöny”. 72 (4–12), s. 348, 1942. (niem.).
- ↑ K. Heißig. Rhinocerotidae aus dem jungtertiär Anatoliens. „Geologisches Jahrbuch”. Reihe B. 15, s. 145–151, 1975. (niem.).
- ↑ von Brandt 1878 ↓, s. 30.
- ↑ Е.И. Беляева. Некоторые данные по олигоценовым носорогам Монголии. „Труды Палеонтологического института”. 55, s. 192, 195, 1954. (ros.).
- ↑ J.B. Hatcher. A median horned Rhinoceros from the loup fork beds of Nebraska. „The American geologist”. 13 (3), s. 149, 1894. (ang.).
- ↑ C.B. Hanson: Teletaceras radinskyi, a new primitive rhinocerotid from the Late Eocene Clarno Formation of Oregon. W: D.R. Prothero & R.M. Schoch (redaktorzy): The evolution of perissodactyls. New York: Oxford University Press, 1989, s. 380, seria: Oxford Monographs on Geology and Geophysics, 15. ISBN 978-0195060393. (ang.).
- ↑ F.A. Lucas. A new Rhinoceros, Trigonias osborni, from the Miocene of South Dakota. „Proceedings of the United States National Museum”. 23 (1207), s. 221, 1901. (ang.).
- ↑ L.T. Holbrook & S.G. Lucas. A new genus of rhinocerotoid from the Eocene of Utah and the status of North American „Forstercooperia”. „Journal of Vertebrate Paleontology”. 17 (2), s. 384, 1997. DOI: 10.2307/4523815. (ang.).
- ↑ D. Geraads, M.L. McCrossin & B. Benefit. A new rhinoceros, Victoriaceros kenyensis gen. et sp. nov., and other Perissodactyla from the Middle Miocene of Maboko, Kenya. „Journal of Mammalian Evolution”. 19 (1), s. 58, 2012. DOI: 10.1007/s10914-011-9183-9. (ang.).
- ↑ Prothero 2005 ↓, s. 124.
- ↑ D.E. Wilson & D.M. Reeder (redaktorzy): Family Rhinocerotidae. [w:] Mammal Species of the World. A Taxonomic and Geographic Reference (Wyd. 3) [on-line]. Johns Hopkins University Press, 2005. [dostęp 2021-05-15]. (ang.).
Bibliografia
- J.F. von Brandt. Tentamen synopseos Rhinooerotidum viventium et fossilium. „Mémoires de l’Académie Impériale des Sciences de Saint-Pétersbourg”. VIIe série. 26 (5), s. 1–66, 1878. (łac.).
- H.F. Osborn. Phylogeny of the rhinoceroses of Europe. „Bulletin of the American Museum of Natural History”. 13 (19), s. 229–267, 1900. (ang.).
- O. Abel. Kritische Untersuchungen über die paläogenen Rhinoeerotiden Europas. „Abhandlungen der Kaiserlich-Königlichen Geologischen Reichsanstalt”. 20, s. 1–52, 1910. (niem.).
- T. Ringström. Nashörner der Hipparion-Fauna Nord Chinas. „Palaeontologia Sinica”. Ser. C. 1 (4), s. 1–156, 1924. (niem.).
- K. Heißig. Paläontologische und geologische Untersuchungen im Tertiär von Pakistan 5. Rhinocerotidae (Mamm.) aus den unteren und mittleren Siwalik-Schichten. „Abhandlungen der Bayerischen Akademie der Wissenschaften (mathematisch-naturwissenschaftliche Klasse)”. Neue folge. 152, s. 1–112, 1972. (niem.).
- 波万黄 & 德阎发 / W. Huang & D. Yan. 湖北神农架板齿犀类新材料 / New material of Elasmotherini from Shennongjia, Hubei. „古脊椎动物与古人类 / Vertebrata PalAsiatica”. 21 (3), s. 223–229, 1983. (chiń. • ang.).
- D.R. Prothero & R.M. Schoch: Origin and evolution of the Perissodactyla: summary and synthesis. W: D.R. Prothero & R.M. Schoch (redaktorzy): The evolution of perissodactyls. New York: Oxford University Press, 1989, s. 504–537, seria: Oxford Monographs on Geology and Geophysics, 15. ISBN 978-0195060393. (ang.).
- C. Guérin. La famille des Rhinocerotidae (Mammalia, Perissodactyla): systématique, histoire, évolution, paléoécologie. „Cranium”. 6 (2), s. 3–14, 1989. (fr.).
- H.Н. Каландадзе & А.С. Раутиан: Система млекопитающих и историческая зоогеография. W: Филогенетика млекопитающих. Москва: Издательство Московского университета, 1992, s. 44–152. (ros.).
- M.C. McKenna & S.K. Bell: Classification of mammals above the species level. Nowy Jork: Columbia University Press, 1997, s. 1–631. ISBN 978-0-231-11013-6. (ang.).
- D.R. Prothero: The Evolution of North American Rhinoceroses. Cambridge: Cambridge University Press, 2005, s. 1–218. ISBN 978-0-521-83240-3. (ang.).