| Eira barbara[1] | |||||
| (Linnaeus, 1758) | |||||
 Samiec hirary amerykańskiej sfotografowany w południowo-wschodniej Brazylii | |||||
| Systematyka | |||||
| Domena | |||||
|---|---|---|---|---|---|
| Królestwo | |||||
| Typ | |||||
| Podtyp | |||||
| Gromada | |||||
| Podgromada | |||||
| Infragromada | |||||
| Rząd | |||||
| Rodzina | |||||
| Podrodzina | |||||
| Rodzaj |
Eira | ||||
| Gatunek |
hirara amerykańska | ||||
| |||||
| Podgatunki | |||||
| |||||
| Kategoria zagrożenia (CKGZ)[50] | |||||
 | |||||
| Zasięg występowania | |||||
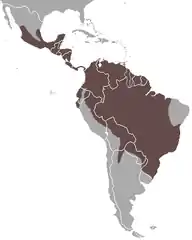 | |||||
Hirara amerykańska[51], hirara (Eira barbara) – gatunek drapieżnego ssaka lądowego z podrodziny Guloninae w obrębie rodziny łasicowatych (Mustelidae). Średniej wielkości ssak drapieżny o długość ciała dochodzącej wraz z ogonem do 117 cm i występujący powszechnie w lasach Ameryki Środkowej i Ameryki Południowej. Jest gatunkiem wszystkożernym, spożywając kręgowce, bezkręgowce, jak i owoce. Samica rodzi od jednego do trzech młodych. Według Międzynarodowej Unii Ochrony Przyrody nie jest zagrożony wyginięciem.
Taksonomia
Gatunek po raz pierwszy zgodnie z zasadami nazewnictwa binominalnego opisał w 1758 roku szwedzki przyrodnik Karol Linneusz, umieszczając go w rodzaju Mustela i nadając mu epitet gatunkowy barbara[8]. Linneusz jako miejsce typowe wskazał łac. Habitat in Brasilia „mieszka w Brazylii”[8], uściślone w 1913 roku przez Einara Lönnberga do Pernambuco w północno-wschodniej Brazylii[40][52][53]. Być może za holotyp posłużył rzeczywisty okaz[19]; Linneusz cytował dzieło „Ac. Holmens” oraz „History of Jamaica” Browne’a (ze znakiem zapytania)[8]; „Galera...The Guinea Fox” Browne’a z „History of Jamaica” opiera się na zwierzęciu „często sprowadzanym na Jamajkę z wybrzeży Gwinei, gdzie jest rodzimym”[54] (przy czym Browne pomylił Gujanę z Gwineą), tym bardziej że w 12. wydaniu „Systema Naturae” Linneusz cytuje tylko Browne’a[55]. Jedyny współczesny przedstawiciel rodzaju hirara[51] (Eira)[56], który nazwał w 1842 roku brytyjski przyrodnik Charles Hamilton Smith[2].
Autorzy Illustrated Checklist of the Mammals of the World rozpoznają siedem podgatunków[57]; jednak analizy przeprowadzone w 2020 roku sugerują, że żaden z wyróżnianych podgatunków nie powinien być uznawany za ważny – podgatunkom opisanym na podstawie wielkości ciała i koloru futra oraz na analizie mtDNA brakuje izolacji geograficznej, a w analizach okazów E. barbara z trzech odrębnych regionów geograficznych w obrębie (domniemanego) ciągłego występowania gatunku zabrakło rozpoznawalnych cech dla wielkości ciała i ubarwienia futra, dla mtDNA czy morfologii czaszek, które można by przypisać do skupisk populacji w obrębie gatunku. Potrzeba dalszych badań nad ostatecznym ustaleniem systematyki podgatunkowej[58]. Podstawowe dane taksonomiczne podgatunków (oprócz nominatywnego) przedstawia poniższa tabelka:
| Podgatunek | Oryginalna nazwa | Autor i rok opisu | Miejsce typowe | Holotyp |
|---|---|---|---|---|
| E. b. inserta | Tayra barbara inserta | J.A. Allen, 1908 | Uluce, Matagalpa, Nikaragua[34]. | Czaszka (nienaruszona, z wyjątkiem brakujących prawych I1, i1, i2 oraz lewego i prawego m1) i skóra (kompletna, rozdarta w okolicy piersiowej) dorosłego samca, sygn. AMNH 28492, zebrana przez W.B. Richardsona 5 stycznia 1908 roku[34]. |
| E. b. madeirensis | Tayra barbara madeirensis | Lönnberg, 1913 | Humaytá, nad rzeką Madeira, Amazonas, Brazylia[45]. | Stary samiec, sygn. BMNH 8.5.9.17, zebrany przez W. Hoffmanna 25 września 1906 roku[40]. |
| E. b. peruana | Galictis barbara, var. peruana | Tschudi, 1844 | Yurac Yacu, San Martín, Peru[10]. | Opis oparty na „G. barbara Linn” Wagnera z 1841 roku[59][21]. |
| E. b. poliocephala | Viverra poliocephalus | Traill, 1821 | Demerara, Gujana[10]. | Wypchany okaz przywieziony do Anglii przez Ch. Edmontona[12]. |
| E. b. senex | Galictis barbara senex | O. Thomas, 1900 | Hacienda Tortugas, Jalapa, na wysokości około 600 ft (183 m), Veracruz, Meksyk[27][60]. | Samiec, sygn. BMNH 89.12.7.4, zebrany w listopadzie 1888 roku i przekazany do Muzeum Historii Naturalnej w Londynie z muzeum w Meksyku[26][60]. |
| E. b. sinuensis | Mustela sinuensis | Humboldt, 1812 | Rzeka Sinú, Bolivar, Kolumbia[45][10]. | Nieznany. |
Ewolucja
Dwa kopalne gatunki Eira zostały opisane z epoki postplioceńskiej ze stanów Maryland i Wirginia pod nazwami Galera macrodon[61] i G. perdicida[62]. Ten drugi gatunek może odnosić się do rodzaju Mephitis, a pierwszy do Trigonictis na podstawie dodatkowego materiału zebranego z osadów wieku ssaków lądowych w Blancan w Stanach Zjednoczonych, a konkretnie ze stanów Waszytngton, Idaho, Nebraska, Kansas, Teksas, Karolina Północna i Floryda[63]. Trigonictis jest uważany za formę pośrednią pomiędzy Galictis i Eira oraz za przodka obu tych gatunków[63]. Ponadto odnotowano wymarłe, plioceńske gatunki Eira ze wschodniej półkuli, ale nie podano okazów typowych, diagnozy ani konkretnych nazw[64]. Hershkovitz (1972) uważa, że Eira oraz inne endemiczne monotypowe rodzaje łasicowatych (np. Lyncodon i Pteronura) z Ameryki Południowej mogły wyewoluować in situ i przemieścić się na północ jako część wymiany fauny Ameryki Północnej i Południowej przez panamski most lądowy[65]. Natomiast Ray i współpracownicy (1981) sugerują, że Eira może pochodzić z Ameryki Północnej[66].
Drzewo filogenetyczne
Uproszczone drzewo filogenetyczne rodziny Mustelidae z wyróżnioną pozycja E. barbara[67]:
| Mustelidae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Etymologia
- Tayra: port. taira i hiszp. taira „hirara”, od tupi tayra „hirara”[68][69].
- Eira: rodzima nazwa używana przez Indian z Boliwii i Paragwaju na określenie „hirary”[70][71].
- Galera: łac. galerum „czapka”, od galea „chełm”[72].
- barbara: łac. barbarus „okrutny, dziki, barbarzyński”, od gr. βαρβαρος barbaros „barbarzyński, nieokrzesany, brutalny”[70][73].
- inserta: łac. insertus „włożony”[74][37].
- madeirensis: rzeka Madeira, Brazylia[75].
- peruana: Peru[76].
- poliocephala: gr. πολιος polios „szary, blady, siwy”; -κεφαλος -kephalos „-głowy”, od κεφαλη kephalē „głowa”[77].
- senex: łac. senex, senis „starsza osoba”[78]; w aluzji do lokalnej hiszpańskiej nazwy hirary „cabeza de viejo” „głowa starca”, która z kolei odnosi się do szarego futra na głowie hirary[26].
- sinuensis: rzeka Sinú, Kolumbia[9].
Zasięg występowania
Hirara amerykańska występuje od południowej części stanu Veracruz w Meksyku przez całą Amerykę Środkową i od południa Ameryki Południowej do północnej Argentyny, występując na całym obszarze z wyjątkiem wysokich Andów i obszarów caatinga (wschodnia Brazylia)[50]. Jest to najczęstszy drapieżnik średniej wielkości na całym obszarze swojego zasięgu[63]. Najrzadszy jest w Meksyku, natomiast powszechny w Ameryce Środkowej i Południowej[63].
Zasięg występowania w zależności od podgatunku[79][57]:
- E. barbara barbara – Brazylia, Paragwaj i Argentyna.
- E. barbara inserta – Salwador i Honduras do Kostaryki.
- E. barbara madeirensis – wschodni Ekwador, północno-wschodnie Peru i zachodnia Brazylia.
- E. barbara peruana – amazońska część Boliwii i południowo-wschodnie Peru.
- E. barbara poliocephala – wschodnia Wenezuela, Gujana i dolna Amazonka.
- E. barbara senex – Meksyk do Gwatemali.
- E. barbara sinuensis – Panama do Kolumbii, zachodniego Ekwadoru, zachodniej Wenezueli oraz wyspa Trynidad.
Morfologia

Długość ciała (bez ogona) 55,9–71,2 cm, długość ogona 36,5–46 cm, długość ucha 0,3–0,42 cm, długość tylnej stopy 0,9–12,3 cm; masa ciała 2,7–7 kg[79][10]. Nie występuje geograficzna zmienność dotycząca wielkości ciała[10]. Dorosłe samce są o 30% większe niż samice; jednak różnica ta nie jest tak duża, jak w przypadku innych rodzajów łasicowatych[10]. Dorosłe samce są również silniej umięśnione wokół szyi i ramion niż samice[10]. Hirara amerykańska jest dużym i smukłym łasicowatym, z wydłużonymi kończynami i długim, pokrytym włosami ogonem[79]. Futro jest krótkie i gęste, z miękkim, brązowym podszerstkiem; grzbiet, nogi, stopy i ogon są koloru ciemnobrązowego do czarnyego, co vostro kontrastuje z ciemnosiwą, szarobrązową lub bladożółtą głową i szyją[79][10]. U niektórych osobników rzadko występuje ciemnobrązowa głowy i szyja, które są nieco jaśniejsze lub w kolorze części grzbietowej[10]. Spodnia część ciała jest ciemnobrązowa lub czarna, z wyjątkiem jasnej, od bladożółtej do pomarańczowej plamy, często trójkątnej, na piersi i gardle[10]. Oczy są czarne z niebiesko-zielonym połyskiem[10]. Młode osobniki są całkowicie czarne, czasami z białą plamą na gardle, białą głową lub ciemnym pasem środkowym, który rozciąga się do ogona[10]. Wraz z wiekiem na włosach ochronnych pojawiają się białe końcówki, nadając zwierzęciu „oszroniony” wygląd[10]. Albinizm i amelanizm są bardziej powszechnie spotykane u hirary niż u innych łasicowatych[10]. W Gujanie spotykana jest również żółta odmiana[79]. Uszy są małe i zaokrąglone, zbieżne z głową i nie wystają ponad czubek głowy. U hirar amerykańskich występują na twarzy długie, sztywne wibrysy. Ogon jest gęsty i o dwie trzecie dłuższy od reszty ciała[10]. Na przednich i tylnych łapach występują silne pazury, przystosowane do biegania i wspinania się, ale nie do kopania czy pływania[10][79]. Nagie od spodu stopy mają krótkie, zakrzywione pazury, połączone błoną międzypalcową z bliższym końcem poduszek[10]. Ponadto hirara posiada duże, czteropalczaste opuszki podeszwowe, podwójne opuszki nadgarstkowe i śródstopia oraz owłosioną piętę[10]. Palce są nierównej długości; pierwszy jest najkrótszy, drugi i piąty pośrednie i mniej więcej równej długości, a trzeci i czwarty są najdłuższe i również mniej więcej równej długości[10]. W konsekwencji opuszki palców, gdy nie są rozłożone, tworzą mocno zakrzywioną linię wokół przedniego brzegu opuszki podeszwowej[10]. U hirary szczególnie dobrze rozwinięte są wibrysy nadgarstkowe[10].

Gruczoły analne hirary amerykańskiej nie są powiększone, a ich wydzielina, choć nieprzyjemna w zapachu, nie jest szczególnie przykra dla człowieka; najwyraźniej nie jest używana przez hirarę w sytuacjach zagrażających jej życiu[10]. Bezpośrednio od części grzbietowej do odbytu ciągnie się płytki, wgłębiony obszar ograniczony bocznie i ogonowo przez niski grzbiet skóry; struktura ta jest podobna pod względem embriologicznym do głębokiej podogonowej torebki gruczołowej występującej w u gatunków z rodzaju borsuk (Meles)[10]. Obszar krocza pomiędzy odbytem a sromem lub moszną jest skąpo owłosiony lub wręcz nagi[10]. W przeciwieństwie do innych rodzajów łasicowatych kość prącia nie rozgałęzia się na wierzchołku[10]. Hirara amerykańska ma cztery sutki[80].
Wzór zębowy: I C P M = 34[79][63]. U hirary maerykańskiej brak pierwszych zębów przedtrzonowych w szczęce i żuchwie, chociaż u kilku osobników zachowany jest czasami pierwszy górny przedtrzonowiec[79][63].




Zakres słyszalności hirary jest niższy niż u kotowatych (Felidae) i szopowatych (Procyonidae), ale wyższy niż u psowatych (Canidae) i niedźwiedziowatych (Ursidae) – górna granica określona jest na 85 kHz, natomiast górna użyteczna granica na 40 kHz[80]. U hirar reakcja węchowa (ang. flehmen) może być wywołana przez wiele silnie pachnących substancji, ale zazwyczaj jest obserwowany w odpowiedzi na mocz innego osobnika; hirara wtedy podciąga usta, marszczy i cofa nos, unosi głowę i wstrzymuje na chwilę oddech[80].
Ekologia i biologia
Siedlisko i zachowanie
_(7979132375).jpg.webp)
Hirary amerykańskie zamieszkują głównie lasy tropikalne i subtropikalne, włącznie z wtórnymi lasami deszczowymi, lasami galeriowymi, lasach mglistymi i suchymi lasami zaroślowymi[79][80]. Czasami widywane są w ogrodach, sadach, na plantacjach cukrowca lekarskiego i kukurydzy oraz na polach uprawnych[79][80]. Hirary są w stanie polować na łąkach, chociaż większość czasu spędzają na obszarach leśnych[79]. Na obszarach llanos w Wenezueli hirary amerykańskie zwykle można znaleźć wzdłuż lasów galeriowych; nocą mogą przemierzać rozległe łąki, prawdopodobnie przemieszczając się z jednego lasu do drugiego[79][80]. W Veracruz w Meksyku siedlisko hirar na ogół ogranicza się do obszarów leśnych[79]. Występują w lasach deszczowych Mata Atlântica w Brazylii, na obszarze całej Amazonii, w lasach liściastych i zaroślowych pantanalu w Paragwaju i Boliwii oraz w galeriach i zaroślach leśnych oraz na trawiastych sawannach w Argentynie, Boliwii i Paragwaju[79][80]. W Belize nie stwierdzono znaczących preferencji siedliskowych hirary, było to jednak prawdopodobnie spowodowane niskim zagęszczeniem gryzoni w siedliskach leśnych, co zmusiło hirary do spędzania większej ilości czasu na polowaniu na łąkach[80].
Hirara amerykańska prowadzi głównie dzienny tryb życia, ze szczytami aktywności we wczesnych godzinach porannych i późnych popołudniowych[79]. Czasami hirara może być aktywna w nocy po północy, zwłaszcza gdy przebywa w pobliżu siedlisk zamieszkanych przez człowieka; wtedy prowadzi bardziej zmierzchowy tryb życia[79][70]. Odpoczywa w dziuplach drzew lub podziemnych norach wykopanych u ich podstawy[79]. W Kostaryce wejście do nory hirary u podstawy pigwicy właściwej (Manilkara zapota) miało średnicę 12 cm[70]. Ponadto gniazda hirary można znaleźć w rozgałęzieniach drzew[70]. W niewoli hirary wolą wysoko zawieszone budki lęgowe bez materiału do budowy gniazda, który jest z nich wyrzucany, jeśli się w ich znajduje[70].

Hirary amerykańskie spędzają dużą część swojego czasu na żerowaniu lub odpoczynku na drzewach[79]; podczas 25% (15 z 61) bezpośrednich obserwacji hirary były widoczne w lub w pobliżu baldachimu lasu, co sugeruje silną tendencję do nadrzewnego styla życia[80]. Zazwyczaj prowadzą samotniczy tryb życia, ale czasami obserwowane są dorosłe pary i małe grupy składające się z matki i jej młodych[79]. Jeden raport[81] o hirarach żyjących w stadach liczących 20–157 zwierząt jest uważany za błędny[70]. Na obszarze llanos w Wenezueli samica z dwoma młodymi utrzymywała areał osobniczy o powierzchni 2,25 km² wokół legowiska do czasu, gdy młode osiągnęły wiek około trzech miesięcy, po tym czasie jej zasięg powiększył się do prawie 9 km²[79]. Po zwiększeniu powierzchni swojego areału osobniczego i odstawieniu młodych od mleka samica codziennie korzystała z nowych nor, które były oddalone od siebie średnio o 867 m[79]. W Belize hirara przemierzała średnio 6,89 km dziennie[79]. Samica w okresie 13-miesięcznej obserwacji zamieszkiwała areał o powierzchni 16,03 km², areał jednego samca w okresie 10-miesięcznej obserwacji wynosił 24,44 km², a areał innego samca podczas 2-miesięcznej obserwacji wynosił jedynie 2,11 km², jednak tak krótki czas prowadzenia obserwacji za pomocą sygnału radiowego mógł być przyczyną niewielkiego rozmiaru obserwowanego areału osobniczego[79]. Areały osobnicze wszystkich trzech zwierząt pokrywały się w znacznym stopniu, co sugeruje, że hirary spędzają niewiele czasu na zachowaniach terytorialnych lub nie spędzają go w ogóle[79][70]. W południowo-wschodniej Brazylii areał osobniczy samicy wynosił 5,3 km²; nie wykazywała ona preferencji dla żadnego konkretnego typu siedliska w obrębie swojego areału, ale pojawiała się też w lasach wtórnych i na łąkach[79].

Hirary podczas badania nieznanego im otoczenia stoją wyprostowane, wykorzystując swoje długie ciała, zwłaszcza gdy przebywają w gęstej trawie[70]. Równie dobrze poruszają się na ziemi i na drzewach, a do utrzymania równowagi używają ogona[70]. Na ziemi chodzą lub kłusują, a biegają z dużą prędkością[70]. Wspinają się i schodzą z pionowych pni głową do przodu, chwytając pień na przemian przednimi i tylnymi stopami[70]. Na leżących konarach poruszają się tak samo, jak to robią na ziemi[70]. Pomimo posiadania częściowej błony pławnej między palcami osobniki trzymane w niewoli unikają wchodzenia do wody; jednak jedna obserwowana hirara przepłynęła przez staw w pogoni za Mazama, a osobniki wyposażone w obroże telemetryczne wielokrotnie przekraczały dużą rzekę podczas wysokiego i niskiego poziomu wody[70].
Pokarm
Hirara amerykańska jest opurnistycznym wszystkożercą i w skład jej pokarmu wchodzą owoce, miód, małe kręgowce, owady i padlina[79][70]. W Wenezueli w 18 próbkach kału stwierdzono trzy gatunki kręgowców (kolczanik plamisty (Pattonomys semivillosus), drzewiszka (Rhipidomys sp.) i legwan zielony (Iguana iguana)) i cztery gatunki owoców (Genipa americana, Zanthozylum culantrillo, Guazuma lomenlosa i Psychotria anceps); zarówno P. semivillous, jak i G. americana znaleziono w 50% próbek[79]; ponadto wszystkie ofiary pochodziły z siedlisk zamkniętych, co potwierdza tezę, że hirara jest głównie zwierzęciem leśnym[70]. W Belize w 31 próbkach kału stwierdzono obecność czterech gatunków małych ssaków: dydelf czarnouchy (Didelphis marsupialis) (9,6%), ryżniak błotny (Oryzomys palustris) (22,5%), bawełniak szczeciniasty (Sigmodon hispidus) (32,3%) i szczur śniady (Rattus rattus) (29,0%); ponadto w składzie pokarmowym stwierdzono 19,4% drobnych ptaków, 58,0% stawonogów i 67,7% owoców[79]. Podstawowym spożywanym owocem było Calocarpum mammosum; inne spożywane owoce to Cecropia mexicana, Astrocaryum standleyanum i Spondias mombin[79]. W 2016 roku w Brazylii obserwowano hirarę próbującą upolować leniwca trójpalczastego (Bradypus tridactylus)[82] natomiast w 2018 roku w Kostaryce zobserwowano, jak hirara upolowała młodego leniwca pstrego (Bradypus variegatus)[83]. Badania przeprowadzone w 2021 roku wykazały, że hirary polują na tak duże zwierzęta jak mazama szara (Mazama gouazoubira) czy na gatunki z rodzaju pancernik (Dasypus sp.), królaka brazylijskiego (Sylvilagus brasiliensis) czy marmozetę czarną (Leontopithecus chrysopygus) i są w stanie zabijać ofiary o masie ciała nawet dwukrotnie większej od ich masy ciała[84]. W niewoli hirary amerykańskie mogą spożywać grzyby, mleko, miód, chleb, jaja, hamburgery, dojrzałe i zgniłe owoce, napoje alkoholowe, małe i duże ptaki, gryzonie do wielkości nutrii amerykańskiej (Myocastor coypus), gady, ryby, padlinę, mięso i kości kozie, karmę dla psów i kotów w puszkach oraz resztki ze stołu[70]. Ofiara oraz inne pokarmy są wykrywane głównie przez zmysł zapachu, ponieważ hirary mają stosunkowo słaby wzrok[79]. Upolowanie ofiary następuje po uporczywym pościgu; hirara nie podkrada się ani nie czutuje na swoją potencjalną ofiarę[79].
Rozmnażanie
W niewoli aktywność rozrodcza dorosłych hirar amerykańskich przypada na okres dzienny[79]. Jednak na wyspie Barro Colorado w Panamie zaobserwowano, jak dwa samce i jedna samica zaangażowały się w nocne zachowania seksualne[79]. Samce wydają się gotowe do kopulacji przez cały rok i są zdolne do romnażania w wieku 18 miesięcy, natomiast u samic pierwszą ruja występuje w wieku około 22 miesięcy; cykl rozrodczy wynosi około 52 dni u młodych samic i około 94 dni u starszych[79]. Ruja może występować kilka razy w roku i trwa 3-20 dni[79]. Ciąża trwa 63-67 dni, nie zachodzi opóźniona implantacja[79]. Samica rodzi 1–3 młodych, ale najczęściej w miocie są dwa[79]. Na wolności samce po okresie rozrodczym opuszczają samice, które samotnie wychowują młode[79]. W niewoli samice nie tolerowały obecności samców w późniejszych stadiach ciąży, jednak w jednej sytuacji samiec, który pozostał z samicą i jej młodymi, ani nie napastował młodych, ani też nie przyczyniał się do poprawy ich dobrostanu[80]. Młode po urodzaniu są niewidome, pokryte sierścią i ważą około 100 g[79]. Uszy otwierają się im po 27–34 dniach, a oczy po 35–47 dniach życia[79]. Zęby mleczne zaczynają pojawiać się w 36. dniu i całkowicie wyrzynają się w 99. dniu życia; zęby stałe pojawiają się w 115. dniu i wyrastają całkowicie w 224. dniu życia[79]. Młode spożywają mleko, a później stały pokarm do 70 dnia życia i są odstawiane od piersi po okołu 100 dniach[79]. Zostało zdefiniowanych pięć etapów rozwoju, które przedstawia poniższa tabela[79]:
| Etap rozwoju | Cykl trwania | Charakterystyka |
|---|---|---|
| Etap niemowlęcy | 1–50 dzień życia | Młode ssą matkę i nie opuszczają legowiska. |
| Etap raczkowania | 50–75 dzień życia | Młode opuszczają legowisko na krótki czas, a matka oprócz mleka zapewnia im stały pokarm. |
| Etap odstawienia od mleka | 75–100 dzień życia | Młode podejmują wycieczki eksploracyjne pobliże legowiska, z samodzielnym karmieniem się owocami i owadami. |
| Etap przejściowy | 100–200 dzień życia | Młode opuszczają legowisko, są definitywnie odstawiane od piersi i zaczynają polować z matką i samodzielnie zabijać zdobycz. |
| Etap rozporoszenia | 200–300 dzień życia | Więzi rodzinne ulegają rozpadowi, a młode rozchodzą się w różne strony. |
Poza norą młode i matka używają klikających odgłosów, aby utrzymać kontakt ze sobą, jeśli nie znajdują się w zasięgu wzroku[79]. Matki chronią swoje dzieci i trzymają je przy środku ciała, gdy pojawi się zagrożenie[79]. Matki wciąż chronią swoje rosnące młode i mogą również nosić je lub ciągnąć z powrotem do legowiska za szyję lub uszy[79]. Samice hirary wraz z kolejnymi miotami stają się bardziej uważne w stusunku do swoich dzieci[80]. Umiejętność łapania potencjalnej zdobyczy rozwija się powoli i musi ona zostać wyuczona przez młode[79]. W wieku zaledwie trzech miesięcy młode są w stanie podążać za gryzoniami i małymi ptakami, i gryźć je po tylnej części ciała czy kończynach[79]. Poprzez nabywanie doświadczenia poprawiają miejsce ugryzień, co z czasem prowadzi do zabójczych ukąszeń w podstawę czaszki[79]. Matka często przynosi ranną zdobycz swoim młodym i pozwala ją zabić, by w późniejszych okresach nauki wypuszczać ją, aby młode mogły ją złapać a potem samodzielnie zabić[79][80].
Interakcje międzygatunkowe
Nie odnotowano drapieżnictwa na hirarze amerykańskiej przez większe kręgowce[80]. We wschodniej Brazylii na hirarze zgłoszono jednego pasożyta wewnętrznego świdrowca amerykańskiego (Trypanosoma cruzi) i dwie pchły – pchłę kocią Ctenocephalides felis felis i Hechtiella lopesi[85][86]. Na zabitej na drodze hirarze w Calakmul w Campeche w Meksyku stwierdzono występowanie trzech nicieni – Molineus sp., Physalopterinae gen. sp. i Angiostrongylus vasosum[87].
Hirara amerykańska jest popularnym gatunkiem eksponowanym w ogrodach zoologicznych i, jako że łatwo się oswaja, jest często trzymana w niewoli jako zwierzę domowe[79][88]. Hirara była kiedyś używana przez rdzennych mieszkańców Ameryki do zwalczania gryzoni[88]. Ze względu na bliskie sąsiedztwo siedliska hirary z ludzkimi, zwłaszcza terenami rolniczymi, drapieżnik ten powoduje według człowieka pewne szkody na plantacjach[88]. Hirara też od czasu do czasu zjada drób i plądruje pola kukurydzy i cukrowca lekarskiego, ale szkody te są zwykle minimalne[88].
Status zagrożenia i ochrona
W Czerwonej księdze gatunków zagrożonych Międzynarodowej Unii Ochrony Przyrody i Jej Zasobów został zaliczony do kategorii LC (ang. least concern „najmniejszej troski”)[50]. Powszechność hirary amerykańskiej w dużej części Ameryki Środkowej i Południowej może być konsekwencją jej ekologicznej odrębności w porównaniu z większością sympatrycznych drapieżników; preferencje siedliskowe, okresy aktywności i preferencje żywieniowe mogą zmniejszać konkurencję międzygatunkową między E. barbara a innymi drapieżnikami[80]. Mimo że prawdopodobnie gatunek ten jest zagrożony lokalnie w wyniku działalności człowieka, a nawet gdy zagrożenia obecnie zmniejszają jego populację, nadal uważany jest za lokalnie pospolity w całym swoim zasięgu i występuje w różnorodnych siedliskach tak naturalnych jak i zaburzonych[50]. Prawdopodobnie w niektórych częściach Meksyku zasięg hirary amerykańskiej został zmniejszony z powodu niszczenia lasów tropikalnych i rozprzestrzeniania się rolnictwa[50][79]. Gatunek ten jest również przedmiotem polowań i ginie na drogach w wielu krajach Ameryki Południowej[50].
Uwagi
- ↑ Nazwa nieważna, odrzucona do celów nomenklatorycznych decyzją ICZN[3].
- ↑ Nomen nudum.
- ↑ Niepoprawna późniejsza pisownia Eirara Lund, 1839.
- ↑ Zmiana nazwy Mustela barbara Linnaeus, 1758[10]; miejsce typowe: „Gujana, Brazylia i kilka innych części Ameryki Południowej”[11].
- ↑ Miejsce typowe: Brazylia[13]; etymologia: epitet gatunkowy Mustela gulo Linnaeus, 1758 (rosomak tundrowy); łac. przyrostek -ina „należący do, odnoszący się do”[14].
- ↑ Nowa nazwa dla Mustela barbara Linnaeus, 1758; etymologia: łac. canescens, canescentis „szarawy”, od canescere „stać się siwiejącym lub białym”, od canere „być szarym”, od canus „szary”[15].
- ↑ Miejsce typowe: Gujana[17]; samica należąca do królweskiej menażerii w Paryżu[17]; etymologia: rodzima, paragwajska nazwa leïra dla hirary[17].
- ↑ Pierwsze użycie obecnej kombinacji nazw.
- ↑ Miejsace typowe: Pernambuco, Brazylia[19]; opis oparty na prymitywnym rysunku księcia Mauritza Johana von Nassau-Siegena, znajdujący się w Bibliotece Berlińskiej[19]; etymologia: etymologia nieznana, Smith nie podał znaczenia epitetu gatunkowego[20]; być może rodzima nazwa.
- 1 2 3 4 5 6 7 8 Kombinacja nazw.
- ↑ Miejsce typowe: Bogotá, Colombia[22]; holotyp: dorosły osobnik nieokreślonej płci, skóra (oprawiona) i czaszka, sygn. MNCN 390, zebrana J.M. Gutiérreza de Albę w 1873 roku[22]; etymologia: nowołac. bimaculatus „dwucętkowy”, od łac. bi- „podwójny”, od bis „dwukrotny”; maculatus „cętkowany”, od maculare „mieć cętki, plamki”, od macula „cętka, plamka”[23].
- ↑ Miejsce typowe: Calovébora, Veraguas, Panama[27]; holotyp: sygn. BMNH 69.7.19.1, zebrany przez E. Arcé[26]; etymologia: gr. βιος bios „życie”[28]; λογος logos „słowo, dyskusja”[29]; „nadana obecnie nazwa zarówno upamiętnia wspaniałą pracę opublikowaną przez panów Salvina i Godmana [Biologia centrali-americana], jak i dokładnie odnosi się do tego zwierzęcia, ponieważ jest to Galictis barbara z «Biologia», chociaż białogłowa forma meksykańska jest również określona w tej pracy”[26].
- ↑ Miejsce typowe: Caroni, Trynidad, Trynidad i Tobago[30]; holotyp: samica, sygn BMNH 99.2.2.1, zebrany przez Henry’ego Caracciolo[30]; etymologia: Trynidad (późnołac. Trinitas „Trójca Święta”, od łac. trini „troje razem”, od tres „trzy”[31]).
- ↑ Miejsce typowe: Mapiri, na wysokości 800 m, górny bieg rzeki Beni, zachodnia Boliwia[32]; holotyp: samiec, sygn. BMNH 1.1.1.57, zebrany przez P.O. Simonsa 27 sierpnia 1908 roku[32]; etymologia: nowołac. brunneus „brązowy”, od średniowoecznołąc. brunius „brązowy”[33].
- ↑ Bonda, Magdalena, Kolumbia[34]; holotyp: dorosły samiec, skóra (rozdarta) i czaszka, sygn. AMNH 15469, zebrany przez H.H. Smitha 6 czerwca 1899 roku[34]; etymologia: tupijska nazwa eîrara dla hirary[35].
- ↑ Miejsce typowe: Tucumán, Argentyna[39]; holotyp: dorosły samiec (szwy nosowe zamknięte), sygn. BMNH 5.10.29.3[39]; etymologia: Tucumán, Argentyna[39].
- ↑ Miejsce typowe: rzeka de Oro, Manabí, Ekwador[41]; holotyp: młoda samica, skóra (kompletna z małymi dziurami i wyłysiała) i czaszka (nienaruszona z wyjątkiem brakującego lewego i prawego I1, lewego i1 oraz prawego pm3, sygn. AMNH 34269, zebrany przez W.B. Richardsona 10 stycznia 1913 roku[42]; etymologia: łac. senilis „starczy, zniedołężniały” (tj. białowłosy, siwy), od senex, senis „starsza osoba”[43].
- ↑ Kombinacja nazw; nie Galictis barbara, var. peruana Tschudi, 1844[45].
- ↑ Miejsce typowe: Passo Fundo, Rio Grande do Sul, Brazylia[47]; etymologia: prof. dr. Hans Krieg (1888–1970), niemiecki zoolog, anatom, podróżnik, kolekcjoner z tropikalnej Ameryki[48][49].
Przypisy
- ↑ Eira barbara, [w:] Integrated Taxonomic Information System (ang.).
- 1 2 3 Smith 1842 ↓, s. 201.
- ↑ F. Hemming. Opinion 417. Rejection for nomenclatorial purposes of volume 3 (Zoologie) of the work by Lorenz Oken entitled Okens Lehrbuch der Naturgeschichte published in 1815—1816. „Opinions and declarations rendered by the International Commission on Zoological Nomenclature”. 14, s. 1–42, 1956–1957. (ang.).
- ↑ L. Oken: Lehrbuch der Naturgeschichte. T. 3. Cz. 2. Jena: August Schmid, 1816, s. 1052. (niem.).
- ↑ P.W. Lund. Palæontologie. „L'Écho du monde savant et l’Hermès: journal analytique des nouvelles et des cours scientifiques”. 5 (430), s. 245, 1839. (fr.).
- ↑ Gray 1843 ↓, s. xx.
- 1 2 Gray 1843 ↓, s. xx, 67.
- 1 2 3 4 C. Linnaeus: Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Wyd. 10. T. 1. Holmiae: Impensis Direct. Laurentii Salvii, 1758, s. 46. (łac.).
- 1 2 F.W.H.A.F. von Humboldt: Tableau synoptique desm singes de l’Ameriqe. W: F.W.H.A.F. von Humboldt & A. Bonpland (redaktorzy): Recueil d’observations de zoologie et d’anatomie comparée, faites dans l’Océan atlantique, dans l’intérieur du nouveau continent et dans la mer du sud pendant les années 1799, 1800, 1801, 1802 et 1803. T. 1. Paris: Shoell, 1812, s. 348. (fr.).
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 Presley 2000 ↓, s. 1.
- 1 2 A.G. Desmarest: Mammalogie, ou, Description des espèces de mammifères. Cz. 1. Paris: Chez Mme. Veuve Agasse, imprimeur-libraire, 1820, s. 157, seria: Encyclopédie méthodique. (fr.).
- 1 2 T.S. Traill. Account of the Lutra vittata, and of the Viverra poliocephalus. „Memoirs of the Wernerian Natural History Society”. 3, s. 440, 1821. (ang.).
- 1 2 H.R. Schinz: Das Thierreich, eingetheilt nach dem Bau der Thiere als Grundlage ihrer Naturgeschichte und der vergleichenden Anatomie von den Herrn Ritter von Cuvier. Cz. 1: Säugethiere und Vögel. Stuttgart und Tübingen: in der J.G. Cotta’schen Buchhandlung, 1821, s. 209. (niem.).
- ↑ Jaeger 1944 ↓, s. 111.
- ↑ The Key to Scientific Names ↓, canescens [dostęp 2022-11-04].
- ↑ M.H.C. Lichtenstein: Verzeichniss der Doubletten des Zoologischen Museums der Königl. Universität zu Berlin: nebst Beschreibung vieler bisher unbekannter Arten von Säugethieren, Vögeln, Amphibien und Fischen. Berlin: T. Trautwein, 1823, s. 4. (niem.).
- 1 2 3 É. Geoffroy Saint-Hilaire & F. Cuvier: Histoire naturelle des mammifères, avec figures originales, coloriées, dessinées d’après des animaux vivants. T. 5. Paris: Chez A. Belin, 1824, s. ryc. xl i tekst język = fr.
- ↑ É. Geoffroy Saint-Hilaire & F. Cuvier: Histoire naturelle des mammifères, avec figures originales, coloriées, dessinées d’après des animaux vivants. T. 7. Paris: Chez A. Belin, 1842, s. Table generale 4. (fr.).
- 1 2 3 Husson 1978 ↓, s. 296.
- 1 2 Smith 1842 ↓, s. 203.
- 1 2 J.J. von Tschudi. „Archiv für Naturgeschichte”. 10 (1), s. 248, 1844. (niem.).
- 1 2 Cabrera 1912 ↓, s. 26.
- ↑ The Key to Scientific Names ↓, bimaculata [dostęp 2022-11-05].
- ↑ F. Martínez. Notas sobre mamíferos Americanos. „Anales de la Sociedad Española de la Historia Natural”. 2, s. 244, 1873. (hiszp.).
- ↑ A. Nehring. Beiträge zur Kenntniss der Galictis-Arten. „Zoologische Jahrbücher”. 1, s. 186, 1886. (niem.).
- 1 2 3 4 5 6 Thomas 1900 ↓, s. 146.
- 1 2 G.S. Miller & R. Kellogg. List of North American Recent mammals. „Bulletin - United States National Museum”. 205, s. 745, 1955. (ang.).
- ↑ Jaeger 1944 ↓, s. 31.
- ↑ Jaeger 1944 ↓, s. 126.
- 1 2 3 Thomas 1900 ↓, s. 148.
- ↑ The Key to Scientific Names ↓, trinitatis [dostęp 2022-11-03].
- 1 2 3 O. Thomas. New mammals from Peru and Bolivia, with a list of those recorded from the Inambari River, Upper Madre de Dios. „The Annals and Magazine of Natural History”. Seventh series. 7 (38), s. 180, 1901. (ang.).
- ↑ The Key to Scientific Names ↓, brunnea [dostęp 2022-11-05].
- 1 2 3 4 Lawrence 1993 ↓, s. 159.
- 1 2 Allen 1904 ↓, s. 36.
- 1 2 3 4 5 6 Allen 1904 ↓, s. 39.
- 1 2 J.A. Allen. Mammals from Nicaragua. „Bulletin of the American Museum of Natural History”. 24, s. 662, 1908. (ang.).
- ↑ Cabrera 1912 ↓, s. 25.
- 1 2 3 4 Lönnberg 1913 ↓, s. 14.
- 1 2 3 Lönnberg 1913 ↓, s. 19.
- ↑ Lawrence 1993 ↓, s. 160.
- ↑ Lawrence 1993 ↓, s. 160–161.
- ↑ The Key to Scientific Names ↓, senilis [dostęp 2022-11-05].
- ↑ J.A. Allen. New mammals from Colombia and Ecuador. „Bulletin of the American Museum of Natural History”. 32, s. 484, 1913. (ang.).
- 1 2 3 Á. Cabrera. Catalogo del los mamiferos de America del Sur. I (Metatheria – Unguiculata – Carnivora). „Revista del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" e Institutio Nacional de Investigación de las Ciencias Naturales”. 4 (1), s. 1–307, 1958. (hiszp.).
- ↑ W.H. Osgood. Mammals of an expedition across northern Peru. „Publication. Field Museum of Natural History”. Zoological Series. 10 (12), s. 162, 1914. (ang.).
- ↑ F.D. de Ávila-Pires. Introdução à mastozoologia do Brasil meridional. „Revista Brasileira de Zoologia”. 4 (2), s. 124, 1987. DOI: 10.1590/S0101-81751987000200004. (port.).
- 1 2 I. Krumbiegel. Die säugetiere der Südamerika-expeditionen Prof. Dr. Kriegs. „Zoologischer Anzeiger”, s. 94, 1942. (niem.).
- ↑ The Key to Scientific Names ↓, kriegi [dostęp 2022-11-05].
- 1 2 3 4 5 6 A.D. Cuarón, F. Reid, K. Helgen & J.F. González-Maya, Eira barbara, [w:] The IUCN Red List of Threatened Species 2016, wersja 2022-1 [dostęp 2022-11-02] (ang.).
- 1 2 Nazwy zwyczajowe za: W. Cichocki, A. Ważna, J. Cichocki, E. Rajska-Jurgiel, A. Jasiński & W. Bogdanowicz: Polskie nazewnictwo ssaków świata. Warszawa: Muzeum i Instytut Zoologii PAN, 2015, s. 156. ISBN 978-83-88147-15-9. (pol. • ang.).
- ↑ Husson 1978 ↓, s. 295.
- ↑ D.E. Wilson & D.M. Reeder (redaktorzy): Species Eira barbara. [w:] Mammal Species of the World. A Taxonomic and Geographic Reference (Wyd. 3) [on-line]. Johns Hopkins University Press, 2005. [dostęp 2022-11-02].
- ↑ P. Browne: The civil and natural history of Jamaica: in three parts. In three dissertations. The whole illustrated with fifty copper-plates: in which the most curious productions are represented of the natural size, and delineated immediately from the objects. London: Printed for the author, and sold by T. Osborne and J. Shipton in Gray’s-Inn, 1756, s. 485. (ang.).
- ↑ C. Linnaeus: Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Wyd. 12. T. 1. Holmiae: Impensis Direct. Laurentii Salvii, 1766, s. 67. (łac.).
- ↑ D.E. Wilson & D.M. Reeder (redaktorzy): Genus Eira. [w:] Mammal Species of the World. A Taxonomic and Geographic Reference (Wyd. 3) [on-line]. Johns Hopkins University Press, 2005. [dostęp 2022-11-02].
- 1 2 C.J. Burgin, D.E. Wilson, R.A. Mittermeier, A.B. Rylands, T.E. Lacher & W. Sechrest: Illustrated Checklist of the Mammals of the World. Cz. 2: Eulipotyphla to Carnivora. Barcelona: Lynx Edicions, 2020, s. 452. ISBN 978-84-16728-35-0. (ang.).
- ↑ M.I. Schiaffini. Are subspecies (of Eira barbara) real?. „Journal of Mammalogy”. 101 (5), s. 1410–1425, 2020. DOI: 10.1093/jmammal/gyaa105. (ang.).
- ↑ J.A. Wagner: Supplementband. W: J.Ch.D. von Schreber: Die Säugthiere in Abbildungen nach der Natur, mit Beschreibungen. Cz. 2: Die Raubthiere. Erlangen: Expedition des Schreber’schen säugthier- und des Esper’schen Schmetterlingswerkes, 1841, s. 214. (niem.).
- 1 2 J. Arroyo-Cabrales, L. León-Paniagua & N.G. Ruiz. Recent appraisal of the Mexican mammals on deposit at The Natural History Museum (London), United Kingdom. „Special Publications, Museum of Texas Tech University”. 71, s. 773, 2019. (ang.).
- ↑ E.D. Cope. An addition to the vertebrate fauna of the Miocene period, with a synopsis of the extinct Cetacea of the United States. „Proceedings of the Academy of Natural Sciences of Philadelphia”. 19, s. 155, 1868. (ang.).
- ↑ E.D. Cope. Synopsis of the Extinct Mammalia of the Cave formations in the United States, with observations on some Myriapoda pound in and near the same, and on some Extinct Mammals of the caves of Anguilla, W. I., and of other localities. „Proceedings of the American Philosophical Society held at Philadelphia for promoting useful knowledge”. 11, s. 177, 1869. (ang.).
- 1 2 3 4 5 6 Presley 2000 ↓, s. 2.
- ↑ W.B. Scott: A history of land mammals in the Western Hemisphere. New York: Hafner Publishing Company, 1937, s. 516–576. (ang.).
- ↑ P. Hershkovitz: The recent mammals of the Neotropical region: a zoogeographical and ecological review. W: A. Keast, F. Erk & B. Glass (redaktorzy): Evolution, mammals, and southern continents. Albany: State University of New York Press, 1972, s. 311–431. ISBN 978-0873951869. (ang.).
- ↑ C.E. Ray, E. Anderson & S.D. Webb. The Blancan carnivore Trigonictis (Mammalia: Mustelidae) in the eastern United States. „Brimleyana”. 5, s. 1–36, 1981. (ang.).
- ↑ Ch.J. Law. Evolutionary shifts in extant mustelid (Mustelidae: Carnivora) cranial shape, body size and body shape coincide with the Mid-Miocene Climate Transition. „Biology Letters”. 15 (5), s. 20190155, 2019. DOI: 10.1098/rsbl.2019.0155. (ang.).
- ↑ Palmer 1904 ↓, s. 666.
- ↑ tayra. Merriam-Webster. [dostęp 2022-11-03]. (ang.).
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 Presley 2000 ↓, s. 4.
- ↑ Palmer 1904 ↓, s. 252.
- ↑ Palmer 1904 ↓, s. 289.
- ↑ The Key to Scientific Names ↓, barbarus [dostęp 2022-11-03].
- ↑ insert. Online Etymology Dictionary. [dostęp 2022-11-03]. (ang.).
- ↑ The Key to Scientific Names ↓, madeirensis [dostęp 2022-11-03].
- ↑ The Key to Scientific Names ↓, peruana [dostęp 2022-11-03].
- ↑ The Key to Scientific Names ↓, poliocephala [dostęp 2022-11-03].
- ↑ The Key to Scientific Names ↓, senex [dostęp 2022-11-03].
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 S. Larivière, A.P. Jennings: Family Mustelidae (Weasels and relatives). W: D.E. Wilson, R.A. Mittermeier (redaktorzy): Handbook of the Mammals of the World. Cz. 1: Carnivores. Barcelona: Lynx Edicions, 2009, s. 624. ISBN 978-84-96553-49-1. (ang.).
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 Presley 2000 ↓, s. 3.
- ↑ E.R. Alston: Mammalia. W: F.D. Godman & O. Salvin (redaktorzy): Biologia Centrali-Americana. London: R. H. Porter, 1879–1882, s. 79–80. (ang.).
- ↑ T.V. Sobroza, A.L. Gonçalves & L.S. dos Santos. Predation attempt and abnormal coat coloration of the tayra (Eira barbara) in the Brazilian Central Amazon. „Studies on Neotropical Fauna and Environment”. 51 (3), s. 231–234, 2016. DOI: 10.1080/01650521.2016.1227137. (ang.).
- ↑ C. Sáenz-Bolaños, V. Montalvo, E. Carillo & T.K. Fuller. Tayra (Eira barbara) predation of a brown-throated three-toed sloth (Bradypus variegatus) in Costa Rica. „Edentata”. 19, s. 70, 2018. (ang.).
- ↑ F. Grotta-Neto, M.C.H. Mello, R.C. Mello, I.P. Bernardi, E. Carrano & F.C. Passos. The role of tayra (Eira barbara) as predator of medium and large-sized mammals. „Austral Ecology”. 46 (2), s. 329–333, 2021. DOI: 10.1111/aec.12982. (ang.).
- ↑ L.R. Guimaräes. Nota sôbre Siphonaptera e redescrição de Polygenis occidentalis (Almeida Cunha, 1914). „Separata da Revista do Museu Paulista”. 2 (6), s. 215–251, 1940. (port.).
- ↑ L.R. Guimaräes & P.M. Linardi. Hechtiella lopesi sp. n. from São Paulo state, Brazil (Siphonaptera: Rhopalopsyllidae). „Memórias do Instituto Oswaldo Cruz”. 88 (4), s. 547–550, 1993. DOI: 10.1590/S0074-02761993000400008. (ang.).
- ↑ J.A. Panti-May, D.I. Hernández-Mena, H.A. Ruiz-Piña, M.A. Torres-Castro, S.F. Hernández-Betancourt & V.M. Vidal-Martínez. Morphological and Molecular Identification of Nematodes in The Tayra Eira Barbara from Campeche, Mexico. „Helminthologia”. 58 (3), s. 315–322, 2021. DOI: 10.2478/helm-2021-0034. (ang.).
- 1 2 3 4 Ch. Schreffler: Eira barbara tayra. [w:] Animal Diversity Web [on-line]. LSA Museum of Zoology. [dostęp 2022-11-06]. (ang.).
Bibliografia
- Ch.H. Smith: Introduction to Mammalia. W: W. Jardine: The naturalist’s library. Cz. 15: Mammalia. Edinburgh: W. H. Lizars, 1842, s. 73–313. (ang.).
- J.E. Gray: List of the specimens of Mammalia in the collection of the British museum. London: The Trustees, 1843, s. 1–216. (ang.).
- O. Thomas. The geographical races of the tayra (Galictis barbara), with notes on abnormally coloured individuals. „The Annals and Magazine of Natural History”. Seventh series. 5 (25), s. 145–148, 1900. (ang.).
- J.A. Allen. New mammals from Venezuela and Colombia. „Bulletin of the American Museum of Natural History”. 20, s. 327–336, 1904. (ang.).
- T.S. Palmer. Index Generum Mammalium: a List of the Genera and Families of Mammals. „North American Fauna”. 23, s. 1–984, 1904. (ang.).
- Á. Cabrera. El concepto de tipo en zoología y los tipos de mamíferos del Museo de Ciencias Naturales. „Trabajos del Museo de Ciencias Naturales”. Serie zoológica. 1, s. 1–32, 1912. (hiszp.).
- A.J.E. Lönnberg. Mammals from Ecuador and related forms. „Arkiv för zoologi”. 8, s. 1–36, 1913. (ang.).
- Edmund C. Jaeger, Source-book of biological names and terms, wyd. 1, Springfield: Charles C. Thomas, 1944, s. 1-256, OCLC 637083062 (ang.).
- A.M. Husson: The mammals of Suriname. Leiden: E. J. Brill, 1978, s. 1–729. ISBN 90-04-05819-2. (ang.).
- M.A. Lawrence. Catalog of Recent mammal types in the American Museum of Natural History. „Bulletin of the American Museum of Natural History”. 217, s. 1–200, 1993. (ang.).
- S.J. Presley. Eira barbara. „Mammalian Species”. 636, s. 1–6, 2000. DOI: 10.2307/0.636.1. (ang.).
- C. Schreffler: Eira barbara. (On-line), Animal Diversity Web, 2003. [dostęp 2008-06-08]. (ang.).
- Mały słownik zoologiczny: ssaki. Warszawa: Wiedza Powszechna, 1978.
- The Key to Scientific Names, J.A. Jobling (red.), [w:] Birds of the World, S.M. Billerman et al. (red.), Cornell Lab of Ornithology, Ithaca (ang.).