Translacja (łac. translatio „tłumaczenie, przeniesienie”) – w biologii molekularnej proces biosyntezy białek na matrycy mRNA. W jego wyniku dochodzi do ostatecznego przetłumaczenia informacji genetycznej zawartej pierwotnie w sekwencji nukleotydów w DNA na sekwencję aminokwasów w łańcuchu polipeptydowym zgodnie z kodem genetycznym[1].
Translacja jest drugim (po transkrypcji) procesem w biosyntezie białka. Powstawanie łańcucha polipeptydowego sterowane jest przez sekwencję mRNA. Translacja odbywa się w cytoplazmie lub na błonach siateczki śródplazmatycznej szorstkiej. Proces ten jest katalizowany przez rybosom obejmujący podjednostkami przesuwającą się nić mRNA. Rybosomy składają się z dwóch podjednostek, małej i dużej, które są zbudowane z białek i rRNA, a funkcję katalityczną pełnią enzymy (rybozymy) zawarte w dużej podjednostce rybosomu. Translacja na jednej cząsteczce mRNA może być prowadzona przez wiele rybosomów równocześnie. Taki kompleks mRNA związanego z wieloma rybosomami nazywa się polisomem lub polirybosomem.
Fazy procesu
Translacja składa się z czterech faz:
- aktywacji
- inicjacji
- elongacji
- terminacji.
W aktywacji właściwy aminokwas jest dołączany do właściwego tRNA za pomocą wiązania estrowego, powstałego przez reakcję grupy karboksylowej aminokwasu i grupy OH przy końcu 3' tRNA. Taki zespół określa się mianem aminoacylo-tRNA.
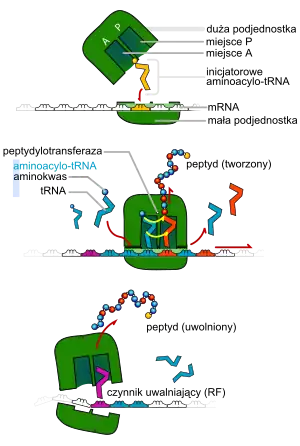
Inicjacja translacji ma miejsce, kiedy mała podjednostka rybosomu przyłącza się do końca 5' mRNA. Do małej podjednostki przyłącza się duża podjednostka rybosomu. Na podjednostce 50s uaktywniają się dwa miejsca: P - miejsce peptydowe i A - miejsce akceptorowe. Pierwszy aminoacylo-tRNA ustawia się w miejscu P.
Elongacja ma miejsce, kiedy następny aminoacylo-tRNA przyłącza się do rybosomu w miejscu A. Następnie proces translacji zachodzi na zasadzie komplementarności kodonu mRNA z antykodonem na tRNA. Rybosom i tRNA są tak ukształtowane, aby dwa aminokwasy, przyłączone do tRNA i zajmujące w rybosomie miejsca A i P, znajdowały się blisko siebie. Dzięki temu zachodzi reakcja między grupą aminową i karboksylową i dwa aminokwasy się łączą. Ten proces – tworzenie wiązań peptydowych – jest katalizowany przez peptydylotransferazę, rybozym (rRNA) wchodzący w skład rybosomu. Po syntezie tRNA szybko zwalnia miejsce P i wraca do cytoplazmy, z kolei aminoacylo-tRNA ulega przesunięciu z miejsca A na miejsce P. Proces ten nazywamy translokacją. Jednocześnie przesuwa się także mRNA. Wielkość tego przesunięcia wynosi zawsze trzy nukleotydy. Na miejsce A nasuwa się nowy aminoacylo-tRNA zawierający antykodon odpowiadający kolejnemu kodonowi na mRNA. Proces elongacji powtarza się aż do napotkania przez podjednostkę mniejszą rybosomu w miejscu A kodonu terminacyjnego (UAA, UAG lub UGA). Tych trójek kodonowych, w normalnych warunkach, nie koduje żaden tRNA.
W tym momencie następuje terminacja translacji. Łańcuch polipeptydowy zostaje uwolniony do cytoplazmy, tRNA zostaje oddzielone od mRNA, a rybosom rozpada się na podjednostki, które mogą zostać ponownie wykorzystane do inicjacji translacji kolejnego mRNA.
Dołączanie kolejnych aminokwasów następuje na końcu C powstającego łańcucha polipeptydowego, a więc translacja zachodzi od końca N do końca C[2].
Translacja u prokariontów
U prokariontów inicjacja translacji wymaga małej i dużej podjednostki rybosomu, czynników inicjacji translacji, GTP (jako źródła energii) oraz inicjatorowego aminoacylo-tRNA (ze związanym aminokwasem formylometioniną). Mała podjednostka rybosomu wiąże się z czynnikiem inicjacji translacji IF3. 16S rRNA z małej podjednostki rybosomu 30S rozpoznaje i wiąże komplementarną sekwencję Shine-Dalgarno w mRNA. Czynnik inicjacji translacji IF2 wiąże się z fMet-tRNA i pomaga mu związać się z małą podjednostką rybosomu. W rybosomie są trzy miejsca, w których może znajdować się tRNA: miejsce A, przez które wchodzi aminoacylo-tRNA (z wyjątkiem pierwszego aminoacylo-tRNA - fMet-tRNA, które wchodzi przez miejsce P), miejsce P, gdzie tworzy się peptydylo-tRNA, oraz miejsce E, przez które tRNA opuszcza rybosom po oddaniu aminokwasu. Aminoacylo-tRNA (fMet-tRNA) znajdujące się w miejscu P rybosomu rozpoznaje kodon inicjujący AUG. Inicjacja kończy się przyłączeniem dużej podjednostki rybosomu i uwolnieniem czynników inicjacji translacji.
Po wejściu fMet-tRNA do miejsca P miejsce A otwiera się i umożliwia związanie się kolejnego aminoacylo-tRNA. W tym wiązaniu bierze udział czynnik elongacji translacji EF-Tu. W ten sposób rozpoczyna się elongacja. Rosnący polipeptyd odłącza się od tRNA w miejscu P, a między ostatnim aminokwasem a aminokwasem przyłączonym do tRNA w miejscu A tworzy się wiązanie peptydowe. Reakcja ta jest katalizowana przez rybozym peptydylotransferazę - 23S rRNA w podjednostce 50S rybosomu. Rybosom przesuwa się o trzy nukleotydy na nici mRNA, z którą są związane tRNA. Dzięki temu polipeptyd przesuwa się z miejsca A do miejsca P, a nienaładowane tRNA trafia do miejsca E. Powstające białko wysuwa się z rybosomu przez otwór w dużej podjednostce. Elongacja trwa, dopóki rybosom nie natrafi na jeden z kodonów terminacyjnych w mRNA.
Kiedy kodon terminacyjny znajdzie się w miejscu A rybosomu, następuje terminacja translacji. Kodony terminacyjne nie są rozpoznawane przez żadne tRNA, tylko przez czynniki uwalniające, które są odpowiedzialne za hydrolizę wiązania estrowego peptydylo—tRNA i uwolnienie nowo powstałego białka z rybosomu. U prokariontów za proces terminacji translacji odpowiedzialne są dwa czynniki — RF1 i RF2. Czynnik RF1 rozpoznaje kodony UAA i UAG, a RF2 rozpoznaje kodony UAA i UGA[3].
Po terminacji mRNA i tRNA są uwalniane z rybosomu, a on sam dysocjuje na podjednostki, które mogą zostać ponownie wykorzystane do inicjacji translacji kolejnego mRNA.
Translacja u eukariontów
U eukariontów inicjacja translacji może przebiegać na dwa sposoby.
Pierwszy z nich to inicjacja zależna od czapeczki. Złożony z czynników inicjacji translacji eIF-4E, eIF4G i eIF4A kompleks eIF4F wiąże się z czapeczką i z białkiem PABP wiążącym ogon poli-A. Mała podjednostka rybosomu 40S wiąże eIF1, eIF3, eIF5 i kompleks eIF2-GTP-Met-tRNAi. Następnie eIF3 wchodzi w interakcje z eIF4G związanym z czapeczką. Taki kompleks inicjacyjny zaczyna przesuwać się wzdłuż cząsteczki mRNA w kierunku od 5' do 3', aż natrafi na kodon start AUG. Wtedy następuje uwolnienie niektórych czynników i związanie się dużej podjednostki rybosomu 60S z małą. Powstaje funkcjonalny rybosom i rozpoczyna się elongacja.
Inicjacja niezależna od czapeczki u eukariontów nie wymaga wędrówki rybosomu od czapeczki do kodonu start. Czynniki ITAF umożliwiają małej podjednostce rybosomu 40S związanie się z sekwencją IRES (ang. internal ribosome entry site - wewnętrzne miejsce wiązania rybosomu). Sekwencje IRES występują m.in. w genach ulegających ekspresji pod wpływem czynników stresowych. Można je znaleźć także u wirusów.
U eukariontów występuje tylko jeden czynnik terminacyjny eRF1[3].
U człowieka o masie 70 kg w ciągu doby syntezowane jest ok. 350 g białek[4].
Translacja a antybiotyki
Różnice w mechanizmach translacji między prokariontami i eukariontami umożliwiają wybiórcze hamowanie syntezy białek bakteryjnych. Antybiotyki zaburzające translację to, między innymi, puromycyna, streptomycyna i inne aminoglikozydy, tetracykliny, chloramfenikol, linkozamidy, makrolidy.
Zobacz też
Przypisy
- ↑ biosynteza białka, [w:] Słownik terminów biologicznych [online], Wydawnictwo Naukowe PWN [dostęp 2020-10-31].
- ↑ Lubert Stryer: Biochemistry; Fifth edition.. W. H. Freeman and Company, 2002, s. 826. ISBN 0-7167-4684-0.
- 1 2 Jan Kutner. Terminacja translacji u Prokaryota i Eukaryota. „Postępy Biochemii”. Tom 53. nr 4, s. 420-430, 2007.
- ↑ Janusz Stanisław Keller: 2.4.7. Anabolizm białek. W: Jan Gawęcki, Lech Hryniewiecki: Żywienie człowieka. T. 1: Podstawy nauki o żywieniu. Warszawa: Wydawnictwo Naukowe PWN, 2003, s. 111. ISBN 83-01-13947-1.