| Cetiosauriscus | |
| von Huene, 1927 | |
| Okres istnienia: kelowej 166–164 mln lat temu | |
 zmontowany holotyp | |
| Systematyka | |
| Domena | |
|---|---|
| Królestwo | |
| Typ | |
| Podtyp | |
| Gromada | |
| Podgromada | |
| Nadrząd | |
| Rząd | |
| Podrząd | |
| Infrarząd | |
| (bez rangi) | Eusauropoda |
| Rodzaj |
cetiozaurysk |
Cetiozaurysk[1] (Cetiosauriscus) – rodzaj wymarłego dinozaura, zauropoda żyjącego między 166 a 164 milionów lat temu (kelowej, jura środkowa) na terenach dzisiejszej Anglii. Był to czworonożny roślinożerca o umiarkowanie – jak na zauropoda – długim ogonie i dłuższych przednich łapach, dorównujących tylnym. Jego długość szacuje się na 15 m, a masę na od 4 do 10 ton.
Jedyna znana skamieniałość obejmuje większość tylnej połowy szkieletu wraz z zadnią kończyną (NHMUK R3078). Znaleziona została w Cambridgeshire w ostatniej dekadzie XIX wieku i opisana przez Woodwarda w 1905 jako nowy okaz gatunku cetiozaura Cetiosaurus leedsi. Jednak w 1927 Friedrich von Huene przeniósł to znalezisko do nowego rodzaju, któremu nadał nazwę Cetiosauriscus, co oznacza „podobny do cetiozaura”. Cetiosauriscus leedsi zaliczony został do zauropodów, a w ich obrębie do rodziny diplodoków – na podstawie podobieństw w budowie ogona i stopy. W 1980 Alan Charig nadał temu gatunkowi nazwę Cetiosauriscus stewarti. Dinozaura tego uznawano początkowo za najstarszego potwierdzonego przedstawiciela diplodoków, ale analiza filogenetyczna z 2003 zaliczyła gatunek do Mamenchisauridae. Kolejne badania z 2005 i 2015 dostarczyły podobnych wniosków – te ostatnie umieściły go poza neozauropodami, choć nie wśród Mamenchisauridae.
Cetiosauriscus znaleziony został w osadach morskich formacji Oxford Clay wśród szczątków wielu różnych grup bezkręgowców, ichtiozaurów, plezjozaurow i krokodyli, a także pojedynczego pterozaura i zróżnicowanych dinozaurów: ankylozaura Sarcolestes, stegozaurów leksowizaura i Loricatosaurus, ornitopoda Callovosaurus, jak też nienazwanych szczątków. Z tej samej formacji znane są skamieniałości teropodów eustreptospondyla, Metriacanthosaurus i megalozaura, jednak prawdopodobnie nie z tego samego poziomu, co cetiozaurysk.
Historia odkryć
Tło

Skamieniałość znana później jako Cetiosauriscus została pierwotnie zaliczona do rodzaju cetiozaur. Ten ostatni należy do pierwszych nazwanych zauropodów (opisał go w 1842 paleontolog Richard Owen), jak również do tych o najbardziej skomplikowanej historii z powodu wielu nieuzasadnionych klasyfikacji znalezisk i gatunków. Mają w niej udział prawie wszystkie angielskie okazy zauropodów. Gatunek typowy cetiozaura zmieniał się z czasem z powodu niekompletności pozostałości i znaczenia taksonu. Wiele aspektów anatomii i pokrewieństw tego rodzaju w dalszym ciągu okrywa niepewność. Cetiosaurus pierwotnie obejmował gatunki C. medius, C. brevis, C. brachyurus i C. longus, żyjące od jury środkowej do kredy wczesnej, pozostawiając po sobie pozostałości w wielu miejscach w Anglii. Żaden z gatunków nie jest jednak w pełni diagnostyczny, a Cetiosaurus jest taksonem ważnym historycznie i taksonomicznie. Bardziej kompletne szczątki należą do środkowojurajskiego gatunku C. oxoniensis nazwanego przez geologa Johna Phillipsa w 1871 i to właśnie on został nowym gatunkiem typowym. W tej samej publikacji autor nazwał również C. glymptonensis, lecz jest on mniej kompletny, a jego pewność pozostawia wątpliwość[2].

Inny angielski takson, Ornithopsis hulkei, nazwany został w 1870 przez paleontologa Harry'ego Seeleya na podstawie kręgów z wczesnokredowej formacji Wessex, co czyni go młodszym od wcześniejszego gatunku cetiozaura[2][3]. Seeley unzał Ornithopsis za blisko spokrewnionego z cetiozaurem, ale odrębnego z uwagi na wewnętrzną budowę kości[3]. Następny gatunek, Ornithopsis leedsii, nazwał w 1887 John Hulke dzięki miednicy, kręgom i żebrom zebranym przez Alfreda Leedsa, angielskiego farmera i amatorskiego łowcę skamieniałości, który zebrał liczne skamieniałości w Oxford Clay[4][5]. O. leedsii żył w jurze późnej. Wykazywał podobieństwa do starszego Cetiosaurus oxoniensis, jako i młodszego O. hulkei[4]. Opisał go bardziej szczegółowo Seeley w 1889, kiedy to zaliczył O. hulkei, C. oxoniensis i O. leedsii do tego samego rodzaju z nazwą Cetiosaurus. Jednak przyrodnik Richard Lydekker podjął z Seeleyem dysputę przed publikacją pracy Seeleya w 1889, sprzeciwiając się uznaniu Cetiosaurus i Ornithopsis za ten sam takson. Lydekker zasugerował, że skamieniałości z Wealden (w tym O. hulkei) należą do Ornithopsis, a szczątki jurajskie (w tym O. leedsii i C. oxoniensis) do Cetiosaurus[6]. Lydekker w 1895 zmienił zdanie i umieścił gatunek O. leedsii w rodzaju pelorozaur. Zaliczano doń już P. brevis, znany wcześniej jako Cetiosaurus brevis). Stworzył więc nową zbitkę P. leedsi, którą z kolei zaliczył do rodziny Atlantosauridae[7]. Popełnił zarazem literówkę, usuwając i z końca epitetu gatunkowego. Powielało ten błąd potem wielu autorów, choć chodzi po prostu o młodszy synonim pierwotnej nazwy[8]. Klasyfikacji Lydekkera nie wsparli późniejsi autorzy, jak paleontolog Arthur Smith Woodward w 1905, który poparł pogląd Seeleya[9].
Odkrycie i nazwa
Skamieniałości znane później jako Cetiosauriscus stewarti odkryli w maju 1898 robotnicy pracujący w glinie w okolicy Fletton na południe od Peterborough i na wschód od Great Northern Railway. Zagłębienia w tym obszarze ujawniły bogate w skamieniałości skały osadowe pochodzenia morskiego zwane Oxford Clay, pochodzące ze środkowego keloweju, dziś uznawane za klasyczną formację geologiczną brytyjskiej paleontologii[10]. Skamieniałości zauropoda prawdopodobnie pochodzą z zagłębienia NPBCL Nr 1, najbardziej na północ wysuniętego wyrobiska, w którym pracowała New Peterborough Brick Company Limited i z którego pochodzi najwięcej skamieniałości kręgowców. Odkrycie przyciągnęło uwagę Leedsa, który, po wydobyciu, zabrał okaz do Eyebury, swego domu rodzinnego. W połowie sierpnia, po oczyszczeniu i innych pracach nad okazem, geolog Henry Woodward odwiedził Eyebury i sporządził naturalnej wielkości rysunki szczątków celem prezentacji dla British Association for the Advancement of Science Meeting. Po prezentacji, 17 sierpnia 1898, Henry Woodward wrócił z amerykańskim paleontologiem Marshem, który uznał zauropoda za blisko spokrewnionego z północnoamerykańskim diplodokiem. Alfred Leeds zaoferował okaz British Museum of Natural History (BMNH, obecnie NHMUK) za £250, co przekłada się na wartość £30529 w 2017[5][11]. NHMUK już wcześniej w 1890 i 1892 kupowało pierwszą i drugą kolekcję Leedsa. Woodward, odpowiadający za zbiory geologiczne muzeum, miał „wielką przyjemność” zarekomendować zarządcom placówki kupno skamieniałości. Handlu dobito 25 lutego 1899, wraz z różnymi innymi szczątkami zakupionymi za £357 (obecna wartość nabywcza £43596[11]). Zauropod Leedsa otrzymał numer BMNH R3078 (obecnie NHMUK R3078)[5].

Zasobność materiału czyni NHMUK R3078 najbardziej kompletnym okazem zauropoda ze Zjednoczonego Królestwa. Porównywalny z nim jest jedynie Dinozaur z Rutland – cetiozaur odkryty w 1967[5][9][12]. Okaz składa się z kończyny przedniej i tylnej oraz kręgosłupa. W przedniej kończynie brakuje ręki, części kości promieniowej i kości łokciowej, podczas gdy w kończynie tylnej nie zachowało się jedynie kilka fragmentów stopy, części kości piszczelowej, strzałkowej i biodrowej. Spośród kręgów poznano cztery fragmenty kręgów grzbietowych, wyrostki kolczyste kości krzyżowej, liczne proksymalne kręgi ogonowe i serię 27 prawie kompletnych kręgów środkowego i dystalnego ogona wraz z połączonymi szewronami. Seria ta nie jest jednak ciągła[5][9]. Koniec ogona (NHMUK R1967) z tej samej lokalizacji pochodzi prawdopodobnie z innego osobnika, a nawet należy do innego taksonu[13].
W 1903 szkielet zmontowano podczas jego pobytu w British Museum, dzięki czemu można go było łatwo porównać z innymi zmontowanymi szkieletami zauropodów, pochodzących z Ameryki Północnej[5][9]. Umieszczono go na wystawie zaraz po odlewie szkieletu diplodoka, wraz z grzbietowymi kręgami NHMUK R1984 i pewnymi izolowanymi zębami kamarazauryda (być może również przypisywanymi Cetiosauriscus[14]). Czyni to cetiozauryska pierwszym zauropodem o szkielecie zmontowanym w Zjednoczonym Królestwie[5].
NHMUK R3078 został w 1905 przypisany przez Arthura Woodwarda do gatunku Cetiosaurus leedsi, jako że pochodził z tej samej formacji geologicznej, co inny okaz umieszczony w rzeczonym gatunku[9]. Woodward zaklasyfikował doń także kręgi grzbietowe NHMUK R1984 i koniec ogona NHMUK R1967[5][9]. W 1927 Huene pokrótce opisał anatomię gatunku C. leedsii, zauważając wiele podobieństw do haplokantozuara. Badacz widział przedmiot swej pracy pomiędzy właściwym cetiozaurem a haplokantozaurem. Dlatego zaproponował dla gatunku nowy rodzaj Cetiosauriscus[15]. Przeniósł do nowego rodzaju NHMUK R1984–R1988 i NHMUK R3078. Huene oparł tworzoną nazwę rodzajową na cetiozaurze. Cetiosaurus oznacza „wielorybi jaszczur”. Dodał do łaciński przyrostek iscus, oznaczający „coś podobnego”. Nazwę rodzajową cetiozaurysk tłumaczy się wobec tego jako „podobny do wielorybiego jaszczura”[16].
Paleontolog Alan Charig w 1980 przypisał wspominany wyżej okaz do rodzaju Cetiosauriscus. Tę klasyfikację NHMUK R1967 uznano za nieprawdopodobną w różnych pracach autorstwa paleontologów takich, jak Friedrich von Huene, Paul Upchurch i Darren Naish. Powodem był brak zachodzenia na siebie fragmentów okazów i niepewna pozycja filogenetyczna[16][17][13][15].
Kolejne gatunki

W późniejszym przeglądzie gadziomiednicznych Huene (1932) przeniósł „Ornithopsis” greppini, nazwany przez siebie w 1922, do rodzaju Cetiosauriscus. Znany materiał, odkryty w kimerydzkich osadach formacji Reuchenette w Szwajcarii, obejmował kręgi grzbietowe i ogonowe, kości kończyny przedniej i tylnej, części stopy – pochodzące od przynajmniej dwóch osobników[18]. Licząca 53 cm długości kość ramienna przypominała budową swój homolog u Cetiosauriscus leedsii. Oba te gatunki pierwotnie odróżniono od rodzaju Cetiosaurus, zwracając uwagę na krótsze kręgi grzbietowe, krótsze przednie kończyny oraz dłuższe tylne[19]. Cetiosauriscus greppini różnią od Cetiosauriscus leedsii ogonowe wyrostki poprzeczne spłaszczone w kierunku przednio-tylnym z wydatnym skrzydłowatym rozszerzeniem do góry, a także tylne blaszki sterczące do dołu z przednich ogonowych wyrostków poprzecznych. Kość krucza o zaokrąglonym obrysie z karbem poniżej stawu ramiennego, bardzo proste trzony kości ramiennej i udowej, dystalny koniec kości ramiennej o kłykciu przyśrodkowym wyższym od bocznego, bardziej proksymalnie umieszczony krętarz czwarty oraz dłuższa i prostsza kość kulszowa. Cechy te zasugerowały odrębność na poziomie rodzajowym, co poparli Upchurch i John Martin w 2003, David Weishampel ze współpracownikami w 2004 i John Whitlock w 2011, który uznał C. greppini za euzauropoda incertae sedis[17][2][20]. Podobieństwa, jak anatomia kręgów ogonowych, zostały użyte przez Christiana Meyera i Basila Thüringa w 2003 do wsparcia umieszczenia tych dwóch gatunków jako odrębnych, ale w tym samym rodzaju[18]. Schwarz i współpracownicy (2020) ustanowili C. greppini gatunkiem typowym odrębnego rodzaju Amanzia[21].
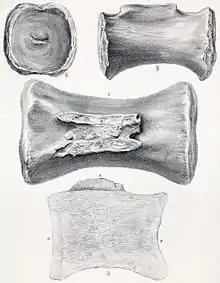
Gatunek Cetiosaurus longus, nazwany w 1842 przez Owena, został przeniesiony do rodzaju Cetiosauriscus bez komentarza przez specjalizującego się w zauropodach paleontologa Johna Stantona McIntosha w 1990[22]. Materiał odnoszony do tego gatunku nazwany został dzięki grzbietowym i ogonowym kręgom z Portland Stone z Garsington, Oxfordshire, dwom innym kręgom ogonowym z tych samych osadów w okolicy Thame i pojedynczego kręgu i kości śródstopia nazwanym wpierw Cetiosaurus epioolithicus (obecnie nomen nudum), chociaż materiał z Garsington został stracony po opisie. Jeden z kręgów (OUMNH J13871) może okazać się szyjnym, jako że ma jedną łagodnie wypukłą i jedną wklęsłą powierzchnię stawową. Diagnozowany przez niezwykle wydłużone trzony kręgów, C. longus nie jest taksonem diagnostycznym. Jako że brakuje mu cech diagnostycznych cetiozauryska, gatunek powinien być nazywany oryginalną zbitką Cetiosaurus longus[2].
Cetiosaurus glymptonensis, nazwany na podstawie dziewięciu środkowo-dalszych trzonów kręgów ogonowych z formacji Forest Marble z angielskiego Oxfordshire, został przeniesiony do rodzaju Cetiosauriscus piórem McIntosha w 1990. Rzeczone kręgi uznano na bardziej wydłużone od należących do Cetiosaurus oxoniensis, jednak proporcja długości kręgów różni się znacznie w obrębie ogona i różnych taksonów, na przykład apatozaur, Diplodocus i Cetiosauriscus cechują się podobnie wydłużonymi kręgami ogonowymi[2]. Bardziej proksymalne kręgi ogonowe mają duży grzbiet w dwóch trzecich górnych trzonu, a mniejszy w jednej trzeciej. Grzbiety te przypominają środkowe kręgi ogonowe cetiozauryska, ale brakuje ich w kręgach ogona tej samej wielkości i proporcji, które z powodu tej różnicy Upchurch i Martin w 2003 przenieśli do innego niż cetiozaurysk rodzaju[2]. „Cetiosaurus” glymptonensis uznawany jest przez nich za Eusauropoda incertae sedis, co popierają Weishampel i współpracownicy (2004) oraz Whitlock (2011). Wymaga odrębnej nazwy rodzajowej, jako że ma pojedynczą cechę diagnostyczną – boczne grzbiety[2][17][20].

W 1980 Charig opisał okaz nieokreślonego bliżej Diplodocidae z wczesnokredowej Anglii, dokonując zarazem powtórnego badania holotypu Cetiosauriscus leedsii celem porównania ich cech. Potwierdził, że kość biodrowa holotypu C. leedsi, NHMUK R1988, była zbyt niekompletna, by ją porównywać z także niekompletną kością biodrową opisywanego okazu NHMUK R3078. Z powodu braku zachodzącego materiału przypisanie NHMUK R3078 gatunkowi Cetiosauriscus leedsii nie było możliwe do potwierdzenia. Charig nazwał nowy gatunek Cetiosauriscus stewarti na podstawie NHMUK R3078. Epitetem gatunkowym uhonorował sir Ronalda Stewarta, szefa London Brick Company, właściciela glinianki, gdzie znaleziono szczątki. Co więcej, Charig uznał Cetiosauriscus leedsii i Cetiosauriscus greppini za nomina dubia, uznając C. stewarti za jedyny pewny gatunek cetiozauryska[8]. Z powodu niepewności gatunku typowego C. leedsii Charig napisał w 1993 petycję do Międzynarodowej Komisji Nomenklatury Zoologicznej, w której wnioskował o uznanie za gatunek typowy dla swego rodzaju Cetiosauriscus stewarti, takson obejmujący okaz nazwany omawianą nazwą rodzajową przez Huene celem odróżnienia do cetiozaura i taksonomicznie bardziej stabilny[16]. W 1995 zaakceptowano to, czyniąc Cetiosauriscus stewarti typem nomenklatorycznym Cetiosauriscus[23]. Jedyny okaz w sposób pewny zaliczany do C. stewarti to jego holotyp NHMUK R3078, ale jest możliwe, że izolowane zęby z Oxford Clay również reprezentują ten takson[5][14].
Budowa

Cetiosauriscus był średniej wielkości zauropodem, czworonożnym zwierzęciem o umiarkowanie długim ogonie i względnie długich przednich kończynach, dzięki czemu ramiona trzymał w poziomie bioder. Jego długość szacuje się na 15 m na podstawie znalezionego szkieletu, porównywalnie z szesnastometrowym cetiozaurem i szesnastoipółmetrowym patagozaurem[24]. Masa Cetiosauriscus jest mniej pewna, zależy od jego miejsca na drzewie filogenetycznym. Rekonstruowany jako diplodoki, został oszacowany przez Paula w 2010[25] na 4 tony. Rekonstruowany jako cetiozaur został oszacowany przez tego samego autora w 2016[24] na 10 ton[24][25].
Kręgi

Kręgi grzbietowe NHMUK R3078 są niekompletne bądź fragmentaryczne. Pojedynczy trzon z przedniej części grzbietu, opistoceliczny, jest mniej więcej tak długi, jak wysoki[9]. Na powierzchniach bocznych widnieją pleurocele po workach powietrznych[15][17]. Pojedynczy trzon ze środka grzbietu jest trochę mniejszy. Pleurocel ma bardziej wydłużony. Również nie zauważa się wklęsłości brzusznej. Znany jest też kręg z tyłu grzbietu, prawdopodobnie ostatni z kręgów przedkrzyżowych. Zachował się jego cały trzon i większość łuku. Jest znacznie skrócony w porównaniu z pozostałymi kręgami grzbietowymi, choć jest mniej więcej tak samo szeroki przez całą swą wysokość. Ma płytki pleurocel, umiejscowiony wyżej na boku trzonu, znikający na łuku kręgowym. W przeciwieństwie do swego proksymalnego odpowiednika jest w niewielkim tylko stopniu opistoceliczny. Wysoki i wąski cienki pionowy grzebień poniżej przedniego wyrostka łuku (hyposphene) zapewniał dodatkowe połączenie stawowe[17], był dobrze wyrażony na łuku. Zachował się także wyrostek kolczysty, spłaszczony i niski, o zwężonym koniuszku, z jedyną dostrzegalną blaszką w postaci spinopostzygapophyseal lamina biegnącej w dół do tylnego roku wyrostka, do tylnych wyrostków stawowych[9]. Nie przypominało to stosunków u większości diplodoków, których wyrostki kolczyste zdobiło wiele blaszek[26]. Zachowały się cztery wyrostki kolczyste kości krzyżowej. Trzy z nich tworzą pojedynczą płytkę kostną, a jedynie czwarty zachowuje swą odrębność, jak i u diplodoka[9][15].
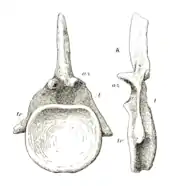
Spośród czterech znanych kręgów ogonowych dwa najbardziej proksymalne są wysoce fragmentaryczne. Każdy z krótkich, ale szerszych niż wyższych trzonów zachowuje ślady wyrostków poprzecznych, stwierdzonych w przypadku innych kręgów. Są one bardzo niskie w porównaniu z kolejnymi kręgami. Bardzo cienkie wyrostki kolczyste robią się jeszcze cieńsze aż do pojedynczego grzebienia z przodu, zwanego prespinal lamina, ale na podobieństwo kręgów grzbietowych noszą po dwie spinopostzygapophyseal laminae. Czwarty z tych kręgów jest najbardziej kompletnym kręgiem proksymalnego ogona. Trzon ma wklęsły z przodu, a płaski z tyłu, a więc amfiplatyczny. W przeciwieństwie do kręgów grzbietowych nie występują pleurocele, a wyrostki poprzeczne zaczynają się w górnej połowie trzonu. Na długość trzon mierzy 10 cm, ma wysokość 27 cm, na szerokość zaś 28 cm. Cały kręg osiąga wysokość 66 cm. Przednie kręgi ogonowe Cetiosaurus leedsi okazu NHMUK R1984 bardzo przypominają te Cetiosauriscus, ale te cetiozauryskowe nie dorównują swym homologom wysokością wyrostków kolczystych, a wyrostkom poprzecznym brak wydatnego grzebienia biegnącego wzdłuż szczytu. Środkowe i dystalne kręgi ogonowe z prawie kompletnej serii 27 kręgów dobrze się zachowały. Te dalsze są nieco bardziej wydłużone od wcześniejszych, jak też nieco bardziej wklęsłe na przedniej powierzchni. W kierunku tylnym trzony stają się coraz mniejsze, a wyrostki poprzeczne również zmniejszają się, by w końcu całkowicie zniknąć, wyrostki kolczyste zaś są coraz krótsze, cieńsze, bardziej pochylone. Siódmy kręg z tej serii przy wysokości 45 cm mierzy 18 cm długości, co czyni go tylko 1 cm dłuższym od kręgu 21 z tej samej serii, osiągającego wysokość 22,5 cm[9]. Cechę wyróżniającą cetiozauryska stanowi obecność wypukłości na szczycie wyrostków kolczystych proksymalnych i środkowych kręgów ogonowych[17].
Kończyny

Zachowała się pojedyncza prawa łopatka, wydłużona i wysmukła. Mierzy 96,5 cm długości i 17,5 cm pośrodku, co czyni ją bardzo wąską[9][19]. Powierzchnia wewnętrzna jest płaska, podczas gdy zewnętrzna – łagodnie wypukła. Nie ma wyrostka na końcu. W pobliżu stawu ramiennego, gdzie łączy się również z kością kruczą, kość staje się cieńsza. Niekompletna kość krucza zachowała się jednak wystarczająco dobrze, by stwierdzić, że jest prostokątna i szersza, niż dłuższa. Kość ramienna mierzy 94 cm długości. Zachowała się w całości z minimalnym tylko zmiażdżeniem. Krótka i korpulentna, nosi wydatną guzowatość naramienną, miejsce przyczepu mięśnia naramiennego, wzdłuż górnej połowy kości[9]. Kształtem kość ramienna przywodzi na myśl skrócenie jej u Neuquensaurus, choć ogólnie kończyna przednia jest długa, jak u Diplodocus i Cetiosaurus, mierząc 69% długości kości udowej[19][17]. Dalszy koniec jest pomarszczony, stanowił miejsce przyczepu dużej chrząstki, podobny stan obserwowano u niektórych innych euzauropodów, jak Amanzia greppini[18]. Kości promieniowa i łokciowa są złamane, ale kompletne. Mierzą 76 cm długości[9].
Kończyna tylna Cetiosauriscus jest o połowę dłuższa od przedniej. Obie kości biodrowe są fragmentaryczne, ale kawałki z przeciwnych stron uzupełniają się wzajemnie, pozwalając na wiarygodny pogląd na proporcje kompletnej kości. Na długość mierzy ona 102 cm. Ma długą i wysmukłą szypułkę łonową[9]. Proporcjonalnie niższa, niż u cetiozaura, przypomina proporcjami Haplocanthosaurus i późniejsze „Titanosauridae”[15][19]. Kompletna lewa kość udowa ma zerodowany trzon. Jest bardzo smukła, mierząc 136 cm wysokości, jednak tylko 19,5 cm szerokości pośrodku[9]. Podobnie smukłą budowę kości udowej dzielił cetiozaurysk z Amphicoelias, Shunosaurus, Ligabuesaurus i jednym z okazów diplodoka, wyprzedzając w tym względzie cetiozaura i większość innych euzauropodów[15][26]. Ma ona wydatny krętarz czwarty, pozostała część trzonu jest jednak bardzo spłaszczona bocznie. Zachowały się także kość piszczelowa, kość strzałkowa i stopa, jednak są one fragmentaryczne i niepołączone stawowo, co utrudnia porównania. Pionowo ustawiona łydka mierzyła jakieś 80 cm[9][19]. Stopa przypomina diplodokową i należącą do Brontosaurus, których pierwszy palec jest duży i zakończony szponem, podczas gdy następne – mniejsze i pozbawione szponów. Najdłuższa jest III kość śródstopia, następne zaś IV, II, V i w końcu I. Ta ostatnia jest z kolei najszersza, szerokość kolejnych kości coraz bardziej maleje[9].
Systematyka

Cetiosauriscus jako rodzaj został pierwotnie zaliczony przez Huene do rodziny cetiozaurydów i podrodziny Cardiodontidae. Podrodzina ta obejmować miała też rodzaje Cetiosaurus, Haplocanthosaurus, Dystrophaeus, Elosaurus i Rhoetosaurus. Oparto ją na ogólnych, pierwotnych cechach wydłużonych kręgów szyjnych i skróconych grzbietowych, w obu przypadkach opistocelicznych, amficelicznych kręgów ogonowych, dystalnie prętowatych, parzystym mostku i kości biodrowej bez wyrostka zapanewkowego (postacetabular process), bardzo szerokiej kości łonowej, szerokiej dystalnej części kości kulszowej, przedniej kończynie znacznie krótszej od tylnej, kości strzałkowej bez przyczepów mięśniowych pośrodku, długich kości śródręcza i krótkich śródstopia[15]. Klasyfikację poprawiono w 1932, kiedy Huene, posiłkując się proporcjami w obrębie przednich i tylnych kończyn, doszedł do wniosku, że Cetiosauriscus jest bliższy Haplocanthosaurus niż cetiozaurowi w obrębie rodziny[19]. Inne stanowisko zajął w 1956, Alfred Romer, który zsynonimizował Cetiosauriscus i Cetiosaurus. Poglądu tego nie potwierdziły kolejne badania taksonu[8][27].
David S. Berman i McIntosh w 1978 umieścili Cetiosauriscus w obfitującej w rodzaje rodzinie diplodoków, wraz z Diplodocus, Apatosaurus, Barosaurus, Mamenchisaurus Dicraeosaurus i Nemegtosaurus. Jak inni jej przedstawiciele, Cetiosauriscus miał skrzydłowate wyrostki poprzeczne, podzielone szewrony z wyrostkami przednimi i tylnymi, biczowaty ogon, kość ramienną długości 2/3 kości udowej, nie miał kości piętowej, z kości śródstopia najdłuższe były III i IV, zaś I miała wyrostek w tylnym dolnym rogu[28]. Klasyfikacja taka czyniłaby znanego z keloweju cetiozauryska najstarszym przedstawicielem tej rodziny, żyjącym miliony lat przed diplodokiem, barozaurem czy apatozaurem. W publikacji nazywającej Cetiosauriscus stewarti Charig opisał też szewrony nowego okazu, na jego określenie tworząc nowe słowo diplodokokształtny („diplodociform”). Oznacza to, że były dobrze zbudowane i miały podwójne szewrony, jak u diplodoka i jego krewnych, do których zaliczano też mamenchizaura. Z powodu podobnych, diplodokokształtnych szewronów Charig przypisał Cetiosauriscus do Diplodocidae wraz z nowym okazem[8]. Pracując nad swym wcześniejszym tekstem, McIntosh (1990[22]) nieśmiało przypisał Cetiosauriscus do podrodziny Diplodocinae, której członkowie cechowali się większą liczbą kręgów szyjnych i mniejszą grzbietowych, wysokimi wyrostkami kolczystymi odcinka krzyżowego, krótkimi przednimi kończynami, brakiem kości piętowej, najdłuższymi III i IV kośćmi śródstopia oraz niewielkim wyrostkiem na dalszym końcu I kości śródstopia. Do podrodziny zaliczały się też Diplodocus, Barosaurus i Apatosaurus[22]. W 2004 pogląd ten poparli bez komentarza Weishampel et al.[17]
Analizę filogenetyczną cetiozauryska przeprowadzili w 2003 Julia Heathcote i Upchurch, bazując na dwóch najpełniejszych matrycach w tym czasie, Wilsona (2002[29]) i Upchurcha (1995[30]). Żadna z nich nie obejmowała wcześniej tego rodzaju. Dodany do analizy Upchurcha Cetiosauriscus okazał się taksonem siostrzanym Tehuelchesaurus w obrębie grupy zawierającej również Mamenchisaurus, Omeisaurus i Euhelopus. Miejsce w grupie z Omeisaurus i Mamenchisaurus znaleziono również przy użyciu matrycy Wilsona. W oparciu o te dwa wyniki Heathcote i Upchurch doszli do wniosku, że Cetiosauriscus nie był nie tylko przedstawicielem diplodoków, ale nawet nie należał do diplodokokształtnych. Zamiast tego był bardziej bazalnym zauropodem poza Neosauropoda[12]. Analiza filogentyczna Rauhuta et al. (2005[31]) pokazała Cetiosauriscus w kladzie z Omeisaurus, klad ten był częścią większej grupy wraz z Losillasaurus i Mamenchisaurus, poza Neosauropoda[31].
Kladogram wedle Rauhut et al. 2005[31] (opisu Brachytrachelopan; uproszczono):
| ||||||||||||||||||||||||||||||||||
Relacje filogenetyczne Cetiosauriscus sprawdzili także w 2015 Tschopp et al. – jako potencjalnego diplodoka. Choć rodzaj zdawał się należeć do Diplodocimorpha wedle jednej z zastosowanych metod, okazał się jednak nie należeć do Neosauropoda. W obu przypadkach Cetiosauriscus stewarti tworzył klad z Barosaurus affinis, wątpliwym gatunkiem znanym jedynie z kości stopy. Tschopp et al. doszli do wniosku, że Cetiosauriscus nie był diplodokiem ani nawet diplodokokształtnym, a wyrzucenie go poza neozauropody zapewnia większa parsymonię niż włączenie we wszystkich analizach do Diplodocoidea. Jako że praca miała za zadanie ustalić pokrewieństwa jedynie wśród Diplodocidae, nie można na jej podstawie poczynić pewniejszych wniosków dotyczących pozycji cetiozauryska[26]. Kladogram wedle Tschopp et al.[26]:

| Eusauropoda |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiologia

Paleopatologia
Seria kręgów ogonowych NHMUK R1967, przypisanych kiedyś cetiozauryskowi, przypomina kręgi ogonowe diplodoka. Są one podwójnie wypukłe, o trzonie długim i cienkim. Obrazują ślady urazu w dwóch miejscach, gdzie widać pozostałości wygojonego złamania[9]. Zmiany zidentyfikowano jako tę samą formę patologii, co znaleziona w ogonie diplodoka[32]. Zasugerowano, że podwójnie wypukłe dalsze kręgi ogonowe zauropodów służyły za wywołujący hałas bicz, cienkie i delikatne nie nadawały się do uderzania. Podatne na urazy stawy czyniły je bezużytecznymi do używania w ten sposób[33].
Paleoekologia

Cetiosauriscus żył w czasach keloweju, piętra epoki jury środkowej, około 166–164 milionów lat temu[34]. Pojedynczy okaz znany jest z Lower Member formacji Oxford Clay wraz z licznymi innymi rodzajami dinozaurów i wielu innych grup zwierząt, z biozony skamieniałości wskaźnikowej Kosmoceras jason[5][14]. Formację Oxford Clay tworzą osady morskie południowej i środkowej Anglii, znane ze świetnej jakości zachowania pewnych skamieniałości i dużej różnorodności taksonów[14]. Osady stanowią ogólnie brązowawoszare mułowce, bogate w materię organiczną obfitą w okruchy pozostałości amonitów i małży, grube na 65 m[35]. Obserwuje się znaczną różnorodność flory, reprezentowaną przez zachowane pyłki i spory. Obecne są nagonasienne i paprotniki, poza tym nieidentyfikowalne kawałki drewna i różnorodny inny materiał roślinny[36].
Z tej samej sekcji formacji geologicznej znany jest zauropod Ornithopsis leedsi, jak również stegozaurydy leksowizaur Lexovisaurus durobrivensis i lorikatozaur Loricatosaurus priscus (być może chodzi o to samo zwierzę), bazalny ankylozaur Sarcolestes leedsi, ornitopod Callovosaurus leedsi i kolejny, nieznany takson ornitopoda. Z dolnej formacji znane są też nieprzypisane węższemu taksonowi dinozaurze jaja. Teropody Eustreptospondylus i być może Megalosaurus także znane są z tej samej formacji, ale pochodzą z osadów nieco młodszych (Middle Member)[17]. Dodatkowo z nieznanego poziomu formacji pochodzi teropod metriakantozaur[14].
Z osadów morskich znane są setki bezkręgowców, w tym małże, ślimaki, łódkonogi, amonity, Teuthoidea, łodzikowca, otwornice, promieniste, mszywioły, ramienionogi, pierścienice, skorupiaki, małżoraczki, wąsonogi i szkarłupnie. Ryby reprezentują Elasmobranchii, Chimaera i Actinopterygii, ichtiozaury – Ophthalmosaurus, plezjozaury – Cryptoclidus, Muraenosaurus, Tricleidus, Liopleurodon, Peloneustes, Pliosaurus i Simolestes, krokodylomorfy – Metriorhynchus i Steneosaurus, pterozaury natomiast ramforynch[14].
Przypisy
- ↑ nazwa polska za: Lambert D., 1996: Księga dinozaurów. Świat Książki, Warszawa, strona 186
- 1 2 3 4 5 6 7 P.M. Upchurch, J. Martin, The anatomy and taxonomy of ''Cetiosaurus'' (Saurischia: Sauropoda) from the Middle Jurassic of England, „Journal of Vertebrate Paleontology”, 23, 2003, s. 208–231, DOI: 10.1671/0272-4634(2003)23[208:taatoc]2.0.co;2.
- 1 2 H.G. Seeley, ''Ornithopsis'', a gigantic animal of the pterodacyle kind from the Wealden, „Annals and Magazine of Natural History”, 28, 5, 1870, s. 279–283, DOI: 10.1080/00222937008696155.
- 1 2 J.W. Hulke, Note on some dinosaurian remains in the collection of A. Leeds, Esq. Part I. ''Ornithopsis Leedsii'', nov. sp, from the Kimmeridge Clay of Northamptonshire, „Geological Magazine”, 8, 4, 1887, s. 375–376, DOI: 10.1017/S0016756800194014.
- 1 2 3 4 5 6 7 8 9 10 L.F. Noè, J.J. Liston, S.D. Chapman, ‘Old bones, dry subject’: the dinosaurs and pterosaur collected by Alfred Nicholson Leeds of Peterborough, England, [w:] R.T.J. Moody i inni, Dinosaurs and Other Extinct Saurians: A Historical Perspective, „Geological Society, London, Special Publications”, 343, 2010, s. 49–77, DOI: 10.1144/SP343.4.
- ↑ H.G. Seeley, Note on the pelvis of ''Ornithopsis'', „Quarterly Journal of the Geological Society”, 1–4, 45, 1889, s. 391–397, DOI: 10.1144/GSL.JGS.1889.045.01-04.27.
- ↑ R. Lydekker, On bones of a sauropodous dinosaur from Madagascar, „Quarterly Journal of the Geological Society of London”, 1–4, 51, 1895, s. 329–336, DOI: 10.1144/GSL.JGS.1895.051.01-04.25.
- 1 2 3 4 A diplodocid sauropod from the Lower Cretaceous of England, [w:] L.L. Jacobs, Aspects of Vertebrate History: Essays in Honor of Edwin Harris Colbert, Museum of Northern Arizona Press, 1980, s. 231–244, ISBN 978-0-897-34053-3.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 A.S. Woodward, On parts of the skeleton of ''Cetiosaurus Leedsi'', a sauropodous dinosaur from the Oxford Clay of Peterborough, „Proceedings of the Zoological Society of London”, January–April, 1, 1905, s. 232–243.
- ↑ J.B. Smith, Oxford Clay, [w:] PJ Currie, K Padian, Encyclopedia of Dinosaurs, 1997, s. 509–510, ISBN 978-0-12-226810-6.
- 1 2 UK Inflation Calculator, CPI Inflation Calculator [dostęp 2018-07-26].
- 1 2 J. Heathcote, P.M. Upchurch, The relationships of ''Cetiosauriscus stewarti'' (Dinosauria; Sauropoda): implications for sauropod phylogeny, „Abstract of Papers, Journal of Vertebrate Paleontology”, Supplement to Number 3, 23, 2003, 60A, DOI: 10.1080/02724634.2003.10010538.
- 1 2 D. Naish, M. Martill, Dinosaurs of Great Britain and the role of the Geological Society of London in their discovery: basal Dinosauria and Saurischia, „Journal of the Geological Society”, 3, 164, 2007, s. 493–510, DOI: 10.1144/0016-76492006-032.
- 1 2 3 4 5 6 D.M. Martill, J.D. Hudson, Fossils of the Oxford Clay, The Palaeontological Association, 1991, s. 1–287, ISBN 978-0-901702-46-3.
- 1 2 3 4 5 6 7 F. von Huene, Sichtung der Grundlagen der jetzigen Kenntnis der Sauropoden, „Eclogae Geologica Helveticae”, 20, 1927, s. 444–470 (niem.).
- 1 2 3 A.J. Charig, Case 2876. ''Cetiosauriscus'' von Huene, 1927 (Reptilia, Sauropodomorpha): designation of ''C. stewarti'' Charig, 1980 as the type species, „Bulletin of Zoological Nomenclature”, 4, 50, 1993, s. 282–283, DOI: 10.5962/bhl.part.1874 [zarchiwizowane z adresu 2018-07-05].
- 1 2 3 4 5 6 7 8 9 DB Weishampel, P Dodson, H Osmólska, The Dinosauria, wyd. 2, University of California Press, 2004, s. 265–784, ISBN 978-0-520-24209-8.
- 1 2 3 C.A. Meyer, C.R. Thuring, Dinosaurs of Switzerland, „Comptes Rendus Palevol”, 1, 2, 2003, s. 103–117, DOI: 10.1016/s1631-0683(03)00005-8.
- 1 2 3 4 5 6 F. von Huene, Die fossile Reptil-Ordnung Saurischia, ihre Entwicklung und Geschichte, „Monographien zur Geologie und Palaeontoiogie”, 4, 1, 1932, s. 1–361 [dostęp 2019-06-11] [zarchiwizowane z adresu 2018-05-11] (niem.).
- 1 2 J.A. Whitlock, The phylogeny of Diplodocoidea (Saurischia: Sauopoda), „Zoological Journal of the Linnean Society”, 4, 161, 2011, s. 872–915, DOI: 10.1111/j.1096-3642.2010.00665.x.
- ↑ Daniela Schwarz, Philip D. Mannion, Oliver Wings i Christian A. Meyer, Re-description of the sauropod dinosaur ''Amanzia ("Ornithopsis/Cetiosauriscus") greppini'' n. gen. and other vertebrate remains from the Kimmeridgian (Late Jurassic) Reuchenette Formation of Moutier, Switzerland, „Swiss Journal of Geosciences”, 113 (1), 2020, s. 2, DOI: 10.1186/s00015-020-00355-5 (ang.).
- 1 2 3 Sauropoda, [w:] David Weishampel, Peter Dodson, Halszka Osmólska, The Dinosauria, University of California Press, 1990, s. 345–401, ISBN 978-0-520-25408-4.
- ↑ International Commission on Zoological Nomenclature, Opinion 1801. ''Cetiosauriscus'' Huene, 1927 (Reptilia, Sauropodomorpha): ''Cetiosauriscus stewarti'' Charig, 1980 designated as type species, „The Bulletin of Zoological Nomenclature”, 52, 1995.
- 1 2 3 G.S. Paul, The Princeton Field Guide to Dinosaurs, wyd. 2, Princeton University Press, 2016, s. 200, ISBN 978-0-691-16766-4.
- 1 2 G.S. Paul, The Princeton Field Guide to Dinosaurs, Princeton University Press, 2010, s. 189, ISBN 978-0-691-13720-9.
- 1 2 3 4 E. Tschopp, O. Mateus, R.B.J. Benson, A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda), „PeerJ”, 3, 2015, e857, DOI: 10.7717/peerj.857, PMID: 25870766, PMCID: PMC4393826.
- ↑ A.S. Romer, Osteology of the reptiles, University of Chicago Press, 1956, s. 620, ISBN 978-0-226-72487-4.
- ↑ D.S. Berman, J.S. McIntosh, Skull and relationships of the Upper Jurassic sauropod ''Apatosaurus'' (Reptilia, Saurischia), „Bulletin of Carnegie Museum of Natural History”, 8, 1978, s. 1–35.
- ↑ J.A. Wilson, Sauropod dinosaur phylogeny: critique and cladistic analysis, „Zoological Journal of the Linnean Society”, 2, 136, 2002, s. 217–276, DOI: 10.1046/j.1096-3642.2002.00029.x.
- ↑ P.M. Upchurch, The evolutionary history of sauropod dinosaurs, „Philosophical Transactions: Biological Sciences”, 349 (1330), 1995, s. 365–390, DOI: 10.1098/rstb.1995.0125, JSTOR: 56238.
- 1 2 3 O.W.M. Rauhut i inni, Discovery of a short-necked sauropod dinosaur from the Late Jurassic period of Patagonia, „Nature”, 7042, 435, 2005, s. 670–672, DOI: 10.1038/nature03623, PMID: 15931221.
- ↑ R.L. Moodie, Studies in paleopathology. I. General consideration of the evidence of pathological conditions found among fossil animals, „Annals of Medical History”, 1, 1917, s. 374–393.
- ↑ N.P. Myhrvold, P.J. Currie, Supersonic sauropods? Tail dynamics in the diplodocids, „Paleobiology”, 4, 23, 1997, s. 393–409, DOI: 10.1017/S0094837300019801.
- ↑ T.R. Holtz, Dinosaurs: The Most Complete, Up-to-date Encyclopedia for Dinosaur Lovers of All Ages, Random House, 2011, Appendix 29, ISBN 978-0-375-82419-7.
- ↑ Peterborough Member, „British Geological Survey”, The BGS Lexicon of Named Rock Units, 2017 [dostęp 2018-08-03].
- ↑ J.B. Riding, A palynological investigation of the Oxford Clay Formation and the Quaternary succession of Northamptonshire (Sheets 171 and 186), „British Geological Survey Internal Report”, IR/04/046, 2004, s. 1–8.
| Wulkanodony | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Euzauropody |
| ||||||||||||||||||||

- kategoria
- klasyfikacja
- lista
- wikiprojekt
| Infrarzędy dinozaurów | |
|---|---|
| Era dinozaurów | |
| Dyscypliny powiązane |
