
Formy przejściowe (pośrednie) – formy organizmów łączących cechy dwóch grup systematycznych.
Występowanie form pośrednich we wcześniejszych epokach geologicznych zakładał Charles Darwin w swoim dziele O powstawaniu gatunków[1]. Twórca teorii ewolucji w wyniku doboru naturalnego pisał:
Ostatecznie, jeżeli moja teoria jest słuszna, to biorąc pod uwagę nie jeden jakikolwiek okres czasu, ale całą przeszłość, musiały bez wątpienia istnieć niezliczone odmiany pośrednie, łączące jak najściślej wszystkie gatunki danej grupy, lecz właśnie sam proces doboru naturalnego dąży nieustannie, jak często wspominaliśmy, do zagłady form rodzicielskich i ogniw pośrednich.
W czasie, gdy powstawało dzieło Darwina, znanych było niewiele skamieniałości wymarłych organizmów. Jednak już w dwa lata po publikacji został odkryty Archaeopteryx, organizm określany często jako pierwszy znany ptak, wykazujący cechy dwóch dużych grup gatunków. W kolejnych latach były odnajdywane skamieniałości, potwierdzające istnienie organizmów o cechach pośrednich między wieloma znanymi współcześnie grupami gatunków[1].
Powszechnym błędem jest przekonanie, że biologia ewolucyjna zajmuje się poszukiwaniem „brakujących ogniw” w historii ewolucyjnej organizmów. Paleontolodzy wykorzystują zapis kopalny przede wszystkim do konstruowania drzew filogenetycznych z uwzględnieniem rozgałęzień powstających w wyniku przemian ewolucyjnych[2]. Pojęcie „brakujące ogniwo” jest wyjątkowo sugestywne i kojarzy się z dotychczas nieodnalezionymi, rzadkimi skamieniałościami form przejściowych, które pozwalają uchwycić początki głównych grup organizmów[3]. Powstanie i rozpowszechnienie pojęcia jest starsze niż teoria ewolucji, nawiązuje do koncepcji wielkiego łańcucha bytu, która porządkuje elementy świata materialnego i metafizycznego w hierarchiczną strukturę określaną jako łańcuch bytu. W XVIII i XIX wieku koncepcja łańcucha bytów została powiązana z myślą ewolucyjną w biologii doprowadzając do jej wypaczenia[2]. Postrzeganie ewolucji jako zmian liniowych od form niższych do wyższych z założeniem postępu zaprzecza idei wspólnego pochodzenia, które opisać można tylko jako szereg rozgałęzień i wzrost różnorodności[4]. Znaczna część „brakujących ogniw” została już dawno odkryta i straciła ten status.
Formy przejściowe a teoria ewolucji
| 1850 |  |
| 1900 |  |
| 1950 | 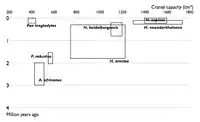 |
| 2002 | 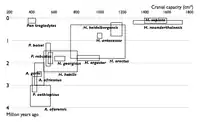 |
| Wykresy pokazujące znane nauce gatunki hominidów w wybranych latach. Każdy gatunek jest pokazany jako prostokąt pokazujący objętość jamy czaszkowej znalezisk danego gatunku oraz daty graniczne znalezisk. Sekwencja diagramów pokazuje wypełnianie przestrzeni pomiędzy uprzednio znanymi gatunkami poprzez nowe znaleziska. | |
W XIX wieku poglądy na ewolucję ukształtowały się w ramach wielkich teorii: darwinizmu, neodarwinizmu, lamarkizmu i neolamarkizmu. Powtórne odkrycie przez H. de Vriea, C. Corensa i E. Tschermaka praw Mendla w roku 1900 (każdy z badaczy dokonał tego niezależnie) doprowadziły do zrozumienia mechanizmów dziedziczenia, rozwoju genetyki i nowego spojrzenia na ewolucję. Lata 1900–1930 zostały określone przez E. Dodsona jako okres agnostycyzmu ewolucyjnego. Dopiero E. Mayr zestawił wiedzę zdobytą w ramach genetyki populacyjnej, ekologii i biogeografii z teorią doboru naturalnego i sformułował teorię specjacji geograficznej, będącą kompletną syntezą wyjaśniającą mikroewolucję. Dowody paleontologiczne nie były dla niej kluczowe[5]. Dane paleontologiczne zostały wykorzystane przez G. Simpsona, który w książce Tempo and Mode in Evolution opublikowanej w roku 1944. Wśród paleontologów teoria Darwina zyskała nielicznych zwolenników. Dominujące były poglądy o charakterze neolamarkizmu i przyjmowanie, że duże grupy systematyczne powstały skokowo. Simpson, będąc paleontologiem, dokonał syntezy wiedzy z zakresu genetyki i paleontologii z teorią darwinowską w zakresie makroewolucji[6]. Używając określania ewolucja kwantowa i wykorzystując model dziedziczenia wielogenowego, Simpson opisał przechodzenie dużych populacji od jednego szczytu przystosowania do kolejnego. Szybkie przejście pomiędzy kolejnymi stanami równowagi jest możliwe przy odpowiednio dużej zmienności populacji i może się dokonać zarówno w wyniku zmian środowiska, jak i zmian wewnątrz organizmu (np. mutacji). Przy zmienności fenotypowej wynoszącej kilka odchyleń standardowych przejście pomiędzy stanami równowagi dokona się przez czas istnienia kilkudziesięciu lub setek pokoleń[7]. W ten sposób zostało wyjaśnione dlaczego formy przejściowe pomiędzy dużymi grupami gatunków zwykle nie zachowują się w postaci skamieniałości[5].
Obecnie teoria ewolucji pozwala na dość precyzyjne ustalenie czasu geologicznego i miejsca, gdzie formy przejściowe powinny się znajdować. Przewidywania te są stosowane na szeroką skalę w praktyce poszukiwawczej paleontologii. Jedną ze skamieniałości odkrytych w ten sposób jest Tiktaalik, forma przejściowa pomiędzy rybami trzonopłetwymi, a wczesnymi czworonogami[8]. Idea istnienia zwierzęcia z płetwami, które wyczołgało się z wody pojawiła się na początku XX wieku. W latach 30. na Grenlandii znaleziono skamieniałości zwierzęcia nazwanego Ichthyostega, pochodzące z późnego dewonu. W literaturze rozpowszechniły się liczne hipotezy i spekulacje związane z przebiegiem ewolucji pomiędzy rybami a zwierzętami czworonożnymi. Dopiero odkrycia z początku XXI wieku pozwoliły przetestować te hipotezy, pokazując, że wiele z nich było fałszywych lub nawet śmiesznych[9].
Przypisy
- 1 2 Sid Perkins. Step-hyphen;'s By-hyphen;'s Step Evolution: Mining the Gaps: Transitional fossils are the hardest to find, but sometimes tell the best stories. „Science News”. 175 (3), s. 30–33, 2009. DOI: 10.1002/scin.2009.5591750320. ISSN 0036-8423. (ang.).
- 1 2 Louise S. Mead. Transforming Our Thinking about Transitional Forms. „Evolution: Education and Outreach”. 2 (2), s. 310–314, 2009. DOI: 10.1007/s12052-009-0126-3. ISSN 1936-6426. (ang.).
- ↑ Per Erik Ahlberg, Jennifer A. Clack. Palaeontology: A firm step from water to land. „Nature”. 440 (7085), s. 747–749, 2006. DOI: 10.1038/440747a. ISSN 0028-0836. (ang.).
- ↑ Emanuele Rigato, Alessandro Minelli. The great chain of being is still here. „Evolution: Education and Outreach”. 6 (1), s. 18, 2013. DOI: 10.1186/1936-6434-6-18. ISSN 1936-6434. (ang.).
- 1 2 Urbanek Jerzy. Biologia XX wieku – główne nurty rozwoju. „Kosmos”. 49 (3 (248)), s. 305–319, 200.
- ↑ Ernst Mayr, William B. Provine: The Evolutionary Synthesis: Perspectives on the Unification of Biology. Harvard University Press, 1998, s. 153–173. ISBN 978-0-674-27226-2. (ang.).
- ↑ Mark Kirkpatrick. Quantum Evolution and Punctuated Equilibria in Continuous Genetic Characters. „The American Naturalist”. 119 (6), s. 833–848, 1982.
- ↑ Edward B. Daeschler, Neil H. Shubin, Farish A. Jenkins. A Devonian tetrapod-like fish and the evolution of the tetrapod body plan. „Nature”. 440 (7085), s. 757–763, 2006. DOI: 10.1038/nature04639. ISSN 0028-0836. (ang.).
- ↑ Jennifer A. Clack. The Fish–Tetrapod Transition: New Fossils and Interpretations. „Evolution: Education and Outreach”. 2 (2), s. 213–223, 2009. DOI: 10.1007/s12052-009-0119-2. ISSN 1936-6426. (ang.).
- kategoria
| Procesy ewolucyjne | |
|---|---|
| Nauki związane z ewolucją | |
| Rodzaje specjacji | |
| Rodzaje doboru naturalnego | |
| Ewolucja molekularna | |
| Ewolucja poszczególnych taksonów | |
| Pojęcia | |
| Historia myśli ewolucyjnej |
|
- Wikiprojekt:Biologia
- kategoria
- wikiprojekt
| Geochronologia | |
|---|---|
| Wymarłe zwierzęta | |
| Rośliny kopalne | |
| Pojęcia |
