
Bakteriorodopsyna – białko o masie 26 kDa występujące u halobakterii należących do domeny archeowców[1]. Zalicza się do białek określanych jako pompy protonowe, posiada zdolność przenoszenia protonów przez błonę komórkową pod wpływem światła[2]. Wytworzona różnica stężeń jonów wodorowych wykorzystywana jest następnie do syntezy ATP. Część naukowców proces syntezy ATP przy udziale bakteriorodopsyny traktuje jako rodzaj fotosyntezy[3]. Sztucznie wytworzone struktury zawierające błony z bakteriorodopsyną oraz syntazą ATP pochodzącą z mitochondriów posłużyły do doświadczalnego potwierdzenia chemiosmiotycznej teorii Mitchella[4]
Bakteriorodopsyna jest integralnym białkiem błonowym zwykle tworzącym dwuwymiarowe plamy, określane jako „purpurowa błona”, których powierzchnia może obejmować do 50% powierzchni komórki archeowców. Powtarzające się elementy sześciokątnej siatki składają się z trzech identycznych łańcuchów białkowych, każdego obróconego o 120 stopni w stosunku do poprzedniego. Każdy łańcuch ma siedem transmembranowych alfa helis i zawiera jedną cząsteczkę retinalu[5].
Trzeciorzędowa struktura bakteriorodopsyny przypomina rodopsynę kręgowców, barwnik odpowiedzialny za reakcję na światło, obecny w siatkówce oka. Pomimo podobnej funkcji obu barwników, sekwencje aminokwasów łańcuchów białkowych znacznie się różnią. Zarówno rodopsyna, jak i bakteriorodopsyna należą do rodziny receptorów białkowych 7TM[6], jednak rodopsyna należy do receptorów sprzężonych z białkiem G, bakteriorodopsyna zaś nie. Struktura bakteriorodopsyny została poznana dzięki rentgenografii strukturalnej i opisana w po raz pierwszy w 1990 roku[7].
Bakteriorodopsyna ma barwię fioletową, a maksimum absorpcji przypada na długości fal odpowiadające barwie zielonej[8] (długość fali 500-650 nm, z maksimum absorpcji w 568 nm).
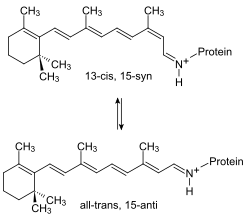
Pochłonięcie fotonu przez cząsteczkę retinalu prowadzi do zmian konformacyjnych części białkowej, których efektem jest przeniesienie protonu przez błonę komórkową. Grupa chromoforowa przyłączona jest kowalencyjnie do Lys216, tworząc zasadę Schiffa[9]. Po fotoizomeryzacji cząsteczki retinalu, proton przenoszony jest na Asp85. Miejsce przyłączenia protonu znajduje się po stronie zewnątrzkomórkowej. Retinal powraca do pierwotnej postaci w wyniku pobrania protonu z Asp96 znajdującej się po stronie wewnątrzkomórkowej. Proton z Asp85 uwalniany jest po stronie zewnątrzkomórkowej, a Asp96 pobiera proton z wnętrza komórki. W efekcie stężenie protonów wzrasta w przestrzeni zewnątrzkomórkowej i maleje wewnątrz komórki[10][11].
Znane są również inne białka o podobnych do bakteriorodopsyny właściwościach, przenosząca jony chlorkowe halorodopsyna oraz aktywowane przez światło kanały jonowe jak ChR1 i ChR2[12].
Aparat fotosyntetyczny niemal wszystkich organizmów fotosyntetyzujących oparty jest na chlorofilu lub bakteriochlorofilu. Pomimo zastosowania innego barwnika mechanizm przekształcania energii świetlnej na energię wiązań chemicznych na halobakterii jest podobny. W pierwszej fazie wytwarzany jest gradient protonowy, a w drugiej energia gradientu wykorzystywana jest przez syntezę ATP. W przypadku chlorofilu możliwe jest wykorzystanie jednocześnie wielu barwników absorbujących światło obecnych w antenach fotosyntetycznych, proces oparty na bakteriorodopsynie umożliwia wykorzystanie jedynie długości fal absorbowanych przez białko. Jest to jedyna forma fotosyntezy obecna u archeowców. Nie jest jednak połączona z asymilacją dwutlenku węgla[13].
Zobacz też
Przypisy
- ↑ Oesterhelt D., Stoeckenius W. Rhodopsin-like protein from the purple membrane of Halobacterium halobium. „Nature: New biology”. 39 (233), s. 149–52, wrzesień 1971. PMID: 4940442.
- ↑ Lanyi JK. Bacteriorhodopsin as a model for proton pumps. „Nature”. 6531 (375), s. 461–3, czerwiec 1995. DOI: 10.1038/375461a0. PMID: 7777054.
- ↑ Lake JA., Clark MW., Henderson E., Fay SP., Oakes M., Scheinman A., Thornber JP., Mah RA. Eubacteria, halobacteria, and the origin of photosynthesis: the photocytes. „Proceedings of the National Academy of Sciences of the United States of America”. 11 (82), s. 3716–20, czerwiec 1985. PMID: 3858845.
- ↑ Racker E., Stoeckenius W. Reconstitution of purple membrane vesicles catalyzing light-driven proton uptake and adenosine triphosphate formation. „The Journal of biological chemistry”. 2 (249), s. 662–3, styczeń 1974. PMID: 4272126.
- ↑ Lanyi JK. Bacteriorhodopsin. „International review of cytology”, s. 161–202, 1999. PMID: 10212980.
- ↑ Varga K., Aslimovska L., Parrot I., Dauvergne MT., Haertlein M., Forsyth VT., Watts A. NMR crystallography: the effect of deuteration on high resolution 13C solid state NMR spectra of a 7-TM protein. „Biochimica et biophysica acta”. 12 (1768), s. 3029–35, grudzień 2007. DOI: 10.1016/j.bbamem.2007.09.023. PMID: 18001693.
- ↑ Henderson R., Baldwin JM., Ceska TA., Zemlin F., Beckmann E., Downing KH. Model for the structure of bacteriorhodopsin based on high-resolution electron cryo-microscopy. „Journal of molecular biology”. 4 (213), s. 899–929, czerwiec 1990. PMID: 2359127.
- ↑ Tittor J., Schweiger U., Oesterhelt D., Bamberg E. Inversion of proton translocation in bacteriorhodopsin mutants D85N, D85T, and D85,96N. „Biophysical journal”. 4 (67), s. 1682–90, październik 1994. DOI: 10.1016/S0006-3495(94)80642-3. PMID: 7819500.
- ↑ Luecke H. Atomic resolution structures of bacteriorhodopsin photocycle intermediates: the role of discrete water molecules in the function of this light-driven ion pump. „Biochimica et biophysica acta”. 1 (1460), s. 133–56, sierpień 2000. PMID: 10984596.
- ↑ Lanyi JK., Lanyi JK. Pathways of proton transfer in the light-driven pump bacteriorhodopsin. „Experientia”. 6-7 (49), s. 514–7, lipiec 1993. PMID: 11536537.
- ↑ Brown LS., Dioumaev AK., Needleman R., Lanyi JK. Connectivity of the retinal Schiff base to Asp85 and Asp96 during the bacteriorhodopsin photocycle: the local-access model. „Biophysical journal”. 3 (75), s. 1455–65, wrzesień 1998. DOI: 10.1016/S0006-3495(98)74064-0. PMID: 9726947.
- ↑ Nagel G., Szellas T., Kateriya S., Adeishvili N., Hegemann P., Bamberg E. Channelrhodopsins: directly light-gated cation channels. „Biochemical Society transactions”. Pt 4 (33), s. 863–6, sierpień 2005. DOI: 10.1042/BST0330863. PMID: 16042615.
- ↑ Lanyi JK. Light energy conversion in Halobacterium halobium. „Microbiological reviews”. 4 (42), s. 682–706, grudzień 1978. PMID: 368557.