| Rya Formation | |
|---|---|
| Stratigraphic range: Early Sinemurian-late Aalenian ~ | |
| Type | Formation |
| Sub-units | Döshult, Pankarp, Katslösa & Rydebäck Members |
| Underlies | Vilhelmsfält & Mariedal Formations |
| Overlies | Höganäs Formation |
| Thickness | Up to 295 m (968 ft) |
| Lithology | |
| Primary | Siltstone, claystone, sandstone, mudstone |
| Other | Coal |
| Location | |
| Coordinates | 56°00′N 12°48′E / 56.0°N 12.8°E |
| Approximate paleocoordinates | 45°24′N 16°42′E / 45.4°N 16.7°E |
| Region | Skåne County Øresund Holstein |
| Country | |
| Extent | Höganäs & Øresund Basins |
| Type section | |
| Named for | Rya, Katslösa |
 Rya Formation (Sweden) | |
The Rya Formation (Swedish: Ryaformationen) is a geologic formation in Skåne County, southern Sweden. It is Early to early Middle Jurassic (early Sinemurian to late Aalenian) in age. The Rya Formation comprises siltstones, claystones, sandstones, mudstones and rare coal beds. The formation overlies the Höganäs Formation and is overlain by the Vilhelmsfält and Mariedal Formations.
The formation was deposited in the Höganäs and Øresund Basins that formed in the earliest Jurassic as part of the break-up of Pangea. The 295 metres (968 ft) thick formation comprises four members, from base to top the Döshult, Pankarp, Katslösa and Rydebäck Members. The depositional environment of the formation ranges from continental to open marine.
The Rya Formation has provided fossils of a number of sharks, ammonites, bivalves and ichnofossils. Coalified wood occurs as scattered pieces up to 6 centimetres (2.4 in) long and indeterminate belemnites, echinoids, serpulids, ostracods and nodosariid foraminifera were also recorded in the formation. Iron ooids containing erratic boulders, called Geschiebe in German, attributed to the Rya Formation were found in Holstein, northern Germany.
Description
The Rya Formation crops out in Colonus Shale Trough of western Scania.[1] The formation overlies the Höganäs Formation and is overlain by the Vilhelmsfält Formation in the Helsingborg area and by the Mariedal Formation in the area of Landskrona and Kävlinge.[2] The Rya Formation is subdivided, from base to top, into the Döshult, Pankarp, Katslösa and Rydebäck Members. The formation is found in the Ängelholm, Helsingborg, Landskrona and Kävlinge areas. In southwest Skåne, the Rya Formation is missing or only poorly developed.[3] In the Øresund Basin between Sweden and Denmark, the formation is truncated by the Basal Middle Jurassic unconformity.[4]
Erratic boulders, called Geschiebe in German, attributed to the Rya Formation and containing iron ooids were found in Holstein, northern Germany.[5]
Subdivision
- Döshult Member
The early Sinemurian Döshult Member (Swedish Döshultsledet) comprises coarse-grained cross-layered sandstones and siltstones in the lower part, and is dominated by dark clays and marls rich in marine fossils in the upper part.[6] The member is up to 80 metres (260 ft) thick in the Ängelholm, Helsingborg and Landskrona areas. Presently, the basal part of this member is exposed at three localities in the Helsingborg area. These contain mineralogically and texturally mature, trough cross-bedded sandstones, commonly with herringbone structures showing north and south oriented paleocurrent directions. The occurrence of herringbone structures in well-sorted sand suggests high energy foreshore to subtidal marine depositional conditions for the lower part of the member. In an abandoned quarry in northwest Skåne (Gantofta brickpit in the Helsingborg area, outcrop very limited at present) the upper part of the Döshult Member commences with bioturbated marine nearshore sands, including burrows, as well as abundant marine invertebrate body fossils. This is followed by a bioturbated shelf mudstone with storm-deposited sand and silt intercalations (tempestites). A massive red mudstone with scarce marine body fossils and burrows follows, which is interpreted as having been deposited rapidly, in a low energy but oxidizing environment. The youngest part of the succession comprises siltstones and mudstones, with carbonate-rich beds, deposited in a shallow marine setting.[3] Coaly detritus, muscovite, very small shells and shell fragments, and framboidal pyrite nodules (0.1–0.3 mm in diameter) are characteristic constituents of this member.[7]
- Pankarp Member
The dark mudstones of the Döshult Member are overlain by the Pankarp Member (Swedish Pankarpsledet) with a sharp conglomeratic boundary.[8] The late Sinemurian Pankarp Member has an estimated thickness of up to 70 metres (230 ft) in the subsurface of the Ängelholm, Helsingborg and Landskrona areas. In the Kävlinge area, the thickness is about 20 metres (66 ft) thick. In westernmost Skåne, the member has been observed in small diameter drill cores. There, the member is subdivided into a lower unit of variegated clays and shales, a middle, poorly sorted silty to sandy unit including a coal bed, and an upper monotonous mudstone unit which is silty and rich in organic matter at the base, and reddish–greenish at the top. It presents different coloration probably due to different degrees of oxidation of iron in the claystone.[6] In the uppermost part of one core, the Pankarp Member comprises lenticular bedded heteroliths with Planolites burrows.[3]
- Katslösa Member

The late Sinemurian to early Pliensbachian Katslösa Member (Swedish Katslösaledet) is mainly known from the subsurface in westernmost Skåne, and it has a thickness of 30 to 40 metres (98 to 131 ft) in the Ängelholm, Helsingborg and Landskrona areas. In the Kävlinge area, the thickness is about 75 metres (246 ft). Sedimentological interpretations are mainly based on the results of petrographical studies of museum collections. The Katslösa Member yields a rich marine microfauna and macrofauna, and it is dominated by homogeneous mudstone deposited in a marine low-energy environment. Is composed mostly by marine green, brown and dark gray claystones and sandstones.[6] Thin beds of matrix-rich quartz wackes are common. They are typically mineralogically mature but texturally highly immature with abundant angular sand grains. The matrix comprises organic matter, micrite, mica and clay minerals. In thin section, the sandstones show evidence of intense burrowing, which has obliterated depositional structures. Scattered berthierine ooids, as well as authigenic siderite crystals have been observed in the member.[9]
- Rydebäck Member
The late Pliensbachian to late Aalenian Rydebäck Member (Swedish Rydebäcksledet) is up to 70 metres (230 ft) thick in the Ängelholm, Helsingborg and Landskrona areas. It is only known from subsurface material in westernmost Skåne, and sedimentological analysis is based on observations from two wells (Rydebäck–Fortuna-1 and -4). These layers were deposited in small bands during a sea regression, and consist of marine gray, black, green and reddish brown sand and siltstones.[6] The member comprises a uniform succession of muddy arenites with a rich marine microfauna (mostly foraminifera), and represents deposition in an offshore low-energy environment. The sediments are strongly burrowed, which has caused an effective mixing of sand and mud, resulting in the forming of quartz wackes. The sand is quartz-rich, and grains are typically well rounded. Berthierine ooids are common constituents of the sediment.[9]
Depositional environments

Tethys Sea transgression entailed formation of fossil-bearing marine deposits in Skane, also associated with an increased tectonic activity.[10] Deposition of the Rya Formation began with nearshore coarse clastics, and continued with offshore mudstones with tempestites (the Döshult Member), followed by offshore muddy sediments with a brief non-marine interval (the Pankarp Member), and ended with deposition of open marine low-energy deposits (Katslösa and Rydebäck Members). The marine Rya Formation shows an overall fining-upwards trend, and an up-section bathymetric deepening of the depositional environment. The depositional environment in western Skåne was either physically protected from the storm energy due to basin topography, or deposition in Skåne took place below storm wavebase. Berthierine ooids occur scattered in the Katslösa Member and are increasingly abundant up-section in the Rydebäck Member. There is a possibility that iron ooid formation was promoted by precipitation of iron and silica from volcanic fluids rising up through the substrate, as has been reported from modern marine sediments offshore Indonesia. This hypothesis has emerged with the publication of age data for the volcanic rocks in Skåne, which now appear to be comparable in age to the prominent iron ooid-bearing deposits, i.e. the Rydebäck Member and the Röddinge Formation.[9]
Age
Based on foraminifers, ammonites and ostracods, the Döshult Member is dated to the early Sinemurian, the Pankarp Member to the late Sinemurian, the Katslösa Member to the late Sinemurian to early Pliensbachian and the Rydebäck Member to the late Pliensbachian to late Aalenian.[3]
The formation is time-equivalent with the Röddinge Formation of the Vomb Trough,[2] the Djupadal Formation in central Skane and the Sorthat Formation of Denmark, with which it shares the Spheripollenites–Leptolepidites and Callialasporites–Perinopollenites Zones.[11][12] The formation also correlates with the Fjerritslev Formation of the Danish Basin,[13] and the Gassum Formation of the Øresund Basin.[14] The storm-dominated, hummocky cross-stratified Hasle Formation on Bornholm is contemporaneous with the muddy Katslösa Member of the Rya Formation.[9]
Basin history

- Basement
The basins where the Rya Formation was deposited form part of the Sorgenfrei-Tornquist Zone (STZ) of the Trans-European Suture Zone, the boundary between Baltica to the northeast and Peri-Gondwana to the southwest. The orogeny was active in the Late Ordovician, or approximately 445 million years ago.
At the Carboniferous-Permian boundary around 300 Ma, the area was influenced by the Skagerrak-Centered Large Igneous Province, another large igneous province stretching across the North Sea, the eponymous Skagerrak between Denmark and Sweden and to the northwest up to northern England and Scotland.

- Break-up of Pangea
The basins of southern Sweden and eastern Denmark were formed during the latest Triassic and earliest Jurassic. During this time the Central Atlantic magmatic province (CAMP), with an estimated 11,000,000 square kilometres (4,200,000 sq mi) the largest igneous province in Earth's history, was formed to the present southwest of the Danish-Swedish realm. In the Skåne area, the Central Skåne Volcanic Province was active during the time of deposition of the Rya Formation, commencing around the Sinemurian-Pliensbachian boundary. The earliest magmatism was partly emplaced into and across pre-existing extensional basin structures.[15] The first and the main volcanic phase of this volcanic province occurred in the Early Jurassic (late Sinemurian to Toarcian) at 191–178 Ma.[16] Analysis of the volcanic rocks produced by this Jurassic volcanism suggests a continental Strombolian-type eruptive character close to the oceans of the Early Jurassic.[17] No correlative pyroclastic beds have yet been identified in sedimentary basins surrounding central Skåne.[18]
- Toarcian
During deposition of the Rydebäck Member, the Toarcian turnover happened. This event at the Pliensbachian-Toarcian boundary characterized by widespread anoxic conditions globally, led to the extinction of various groups of flora and fauna. Taxa inhabiting the upper water column were unaffected by anoxia and included ammonites and belemnites. Epifaunal taxa adapted to low-oxygen conditions, such as the buchiids, posidoniids and inoceramids, flourished in the post-extinction environment during the survival interval.[19]

Economic geology
A study on the geothermal potential of reservoirs in the Øresund Basin published in 2018 by Erlström et al. gave results of the formation together with the Gassum and Höganäs Formations, giving the following characteristics of the three Early Jurassic formations:[20]
- Net sand thickness - 60 to 100 metres (200 to 330 ft)
- Porosity - 18 to 34%
- Permeability - 50 to 1500 mD
- Cl− concentration - 120 to 190 gram/liter
- Productivity index - 7.0 m3/hr/bar
A study published in the same year analyzing the CO2 storage potential of the Rya and Höganäs Formations concluded a storage capacity of 543 megatons of carbon dioxide.[21]
The organic content of the Jurassic strata in Skåne is typically dominated by gas-prone kerogen (type III), which is below, or at the onset of, thermal maturity.[18]
Paleoenvironment
The sedimentological evolution of the Jurassic in southwestern Skåne, specially the Rya Formation, has provided depth diverse data about its different environments, specially on those samples recovered on the Höllviken-2 core and sidewall cores from the FFC-1 well.[22] The unit was linked on all its sedimentological history with the main Fennoscandinavian land. The local succession in the Höllviken Halfgraben and Barsebäck Platform has measured continue during most of the Triassic and Early Jurassic times.[23] Inte western part of the Höllviken Halfgraben during the Hettangian–Pliensbachian succession there is evidence of higher subsidence rate along the Öresund Fault, indicating tectonically controlled deposition and the presence of a submarine high at the west of this fault.[24] In the east is the Skarup Platform, interpreted as a Jurassic high based on its incomplete or missing Rhaetian–Lower Jurassic strata, along redeposited spores and pollen in the Höllviken Halfgraben and increased sand output, that point to this zone as the major freshwater terrestrial source.[25][26] This deposits shows several nearshore environments, from offshore marine environment probably below wave base on the sinemurian level, as proven by the presence of well cemented fine sand and 2 m thick clay dominated heterolites, that change to more marine influenced flaser bedded heterolites with some strongly bioturbated horizons and convolute beds in the Pliensbachian, and end in the Pliensbachian–Lower Toarcian with a sandy seashore setting with high energy lenticular beds.[27] Some sections are suggested to be tidal flat zones and/or distal bar deltaic deposits.[28] Coeval with the Rydebäck member, at Anholt (Fjerritslev Formation) where studied several levels that point to a connected fluvial system.[29] Concretely the Upper Pliensbachian-Toarcian boundary marks the beginning of a major regression, which continued through the Toarcian and Aalenian.[29] The discovery of the foraminifera Eoguttuliiia liassica, Bony Fishes and Scolecodonts, points that the Pliens-Toar boundary has a shallow water environment, possibly of reduced salinity.[30] The Pollen and spores, identical of those found on the Rydebäck member (Vilhelmsfält Bore No. 1 and Karindal bore no. 1), increase, and the dinoflagellate cysts decrease, suggesting more proximal shoreline.[30] This layers, on both Anholt and the Rydebäck member, are considered to represent a regression from marine inner shelf environments to near lagoonal or deltaic conditions during the late Pliensbachian and early Toarcian, but with light marine influence, as show the presence of Dactylioceras on the Rydebäck member.[30] The sequence points concretely than in both units The Late Pliensbachian sediments at were deposited in a storm-influenced Inner Shelf setting, that changed on the Lower Toarcian into a more restricted, marginal-marine conditions with delta progradation.[31] Then in the late Toarcian increased brackish conditions, to end on a short marine encroachment during the Early Aalenian.[31]
The study of the Jurassic sediments has allowed to know that the basement rocks of southern Sweden were deeply weathered in Late Triassic-Cretaceous times, with formed saprolites on the sub-mesozoic basement and high amount of Kaolinite on the stronger weathered profiles, opposed to smectite on the less weathered ones.[32] Thus, fluvial currents released smectite-rich weathering material to the Late Triassic–Jurassic receiving basins.[32] The members of the Rya Formation recover a transition from deltaic to paralic coast and shallow marine, dominated by kaolinite, along with peaks of illite and smectite.[33] The record of this minerals showed that at the time of deposition of the Rya Formation, mid-latitude warmth and pronounced humidity to drier pseudomediterranean climates allowed on the denuded bedrock in the Fennoscandian Shield, composed of Gneisses or Granites, at places intersected by Dolerite dykes, fracturation and active erosion/weathering.[34] The coeval Djupadal Formation volcanic ash falls at the east Skarup Platform-Cenntral Skane Volcanic Province, may have diluted the marine sediments, with raised smectite content. The pronounced mineralogical maturity of most Swedish post-Norian Mesozoic arenites confirms widespread feldspar destruction in the weathering profiles of the Paleozoic crystalline basement at the north, as a consequence of the increased humidity.[35] A fish tooth recovered from the lower Toarcian of the North West German Basin presents a radiogenic seawater value of −6 ε-units, which is quite counter-intuitive to the idea of massive unradiogenic crustal-derived inputs from Laurasia. Clastic fractions found on the same layer suggest brief radiogenic Nd influxes from the Skåne flood basalts erupted at this time.[36]
- Pliensbachian
Lower-late Pliensbachian Layers at Katslösa are similar in faunal composition. The Katslösa Member composition suggest deposition on a Low Energy Marginal marine environment with absence of changes in the salinity (As proven by the presence of Echinoderms and other low salt intolerant fauna), with active bottoms filled by traces of diverse invertebrates and abundance of Bivalves.[9] The Upper Pliensbachian section that mark the appearance of the Rydebäck Member continues the stable salinity environment with increased presence of Echinoderms, but lack of a diverse Bivalve fauna as in the older section. This unit was deposited likely on a marine setting more proximal to the shore and low-energy, more strongly burrowed than older layers. The presence of Selachian fauna can be more likely derived from a more suitable depositional setting for this remains than a biota turnover.[37] Palynology in this section is dominated by spores in the Karindal bore no. 1, suggesting a humid climate on nearby emerged lands.[26]
- Toarcian
Toarcian layers of the formation where influenced by the ongoing vulcanism located in the Central Skåne Volcanic Province (Djupadal Formation), as, marine water influence is observed in the main outcrop of the last unit, where enriched Zeolite by Barium is suggested to derive from oceanic water, that may have circulated as hydrothermal flows in the lapilli tuff and facilitating diagenetic changes.[38] Is also known by palynological analysis on the Bonnarp Cone, where saltwater/brackish acritarchs like Leiosphaera and Leiofusa or Dinoflajellates like Nannoceratopsis where recovered.[39] This section also recovers the increased influence of Terrestrial weathering measured in the layers after the Toarcian AOE, as seen in the Vilhelmsfalt borehole, where plant remains increase their presence, suggesting a regression of the coast influenced by the increased volcanic-derived materials.[40] The Palynology in this section is dominated by the Pollen of Chasmatosporites (Cycads), with at least six species recovered, playing more than 50% of the total palynological samples, implicating a clear dominant role of the producer of this Pollen and suggesting along the increased amount of Cheirolepidiaceae pollen and general decrease of spores a shif towards a more arid climate on nearby settings.[41]
Fossil content
The formation has provided fossils of typically marine fauna. With the exception of a continental coal bed, the formation is marine in character. Shark teeth were reported from the Rydebäck Member.[42]
Annelida
Echinodermata
Ammonites
- Agassiceras nodulatum[46]
- Amaltheus margaritatus[47]
- Amauroceras ferrugineum[48]
- Arnioceras cf. falcaries[49]
- Arnioceras sp. indet.[50]
- Asteroceras obtusum[51]
- Coroniceras rejnesi[52]
- Cymbites striaries[53]
- Dactylioceras tenuicostatum[47][54]
- Dactylioceras cf. semicelatum[47]
- Echioceras raricostatum[55]
- Eleganticeras elegantulum[56]
- Eparietites sp. indet.[57]
- Euagassiceras resupinatum[51]
- Euagassiceras spinaries[51]
- Euagassiceras lundgreni[51]
- Euagassiceras cf. lundgreni[51]
- Megarietites meridionalis[58]
- Oxynoticeras oxynotum[55]
- Oxynoticeras? sp. indet.[59]
- Paracoroniceras crossi[60]
- Pleuroceras spinatum[47]
- Pleuroceras hawskerense[61]
- Polymorphites angustus[62]
- Prodactylioceras davoei[55]
- Promicroceras planicostatum[51]
- Promicroceras sp. juv.[63]
- Tragophylloceras ibex[55]
- Uptonia jamesoni[47]
Belemnites
- "Belemnites" elongatus[64]
- "Belemnites" milleri[65]
- Passaloteuthis alveolata[66]
- Passaloteuthis apicicurvata[66]
- Passaloteuthis cf. virgata[66]
- ?Passaloteuthis sp.[67]
- Pseudohastites charmouthensis[66]
- Pseudohastites cf. arundineus[66]
Brachiopods
- Rhynchonella deffneri[68]
- Spiriferina walcotti[68]
- Zeilleria numismalis[68]
- Zeilleria perforata[68]
Bivalves
- Anomia pellucida[69]
- Anisocardia luggudensis[70]
- Arcomya decora[71]
- Arcomya cf. elongata[72]
- Astarte angelini[73]
- Astarte fructuum[74]
- Astarte scanensis[74]
- Astarte fortuna[75]
- Astarte deltoidea[76]
- Astarte oerbyensis[77]
- Astarte ryensis[77]
- Astarte sp.[7]
- Barbatia pulla[78]
- Cardinia tollini[79]
- Cardinia expansa[80]
- Cardinia ingelensis[81]
- Cardinia kullensis[82]
- Cardinia spp.[7]
- Chlamys interpunctata[7]
- Chlamys textoria[7]
- Chlamys janiformis[83]
- Chlamys subulata[84]
- Chlamys tullbergi[85]
- Chlamys textaria[85]
- Chlamys interpunctata[86]
- Dimyodon sp.[87]
- Entolium sp.[7]
- Entolium hehli[88]
- Entolium calvum[88]
- Entolium cingulatum[89]
- Entolium lundgreni[90]
- Eotrapezium gerrnari[91]
- Eotrapezium pullastra[91]
- Eotrapezium heberti[92]
- Eotrapezium menkei[70]
- Gervillia angelini[93]
- Gervillia scanica[94]
- Gervillia hagenowi[95]
- Gervillia sjögreni[96]
- Goniomya heteropleura[97]
- Grammatodon cypriniformis[98]
- Grammatodon sinuatus[99]
- Grammatodon subrhomboidalis[100]
- Homomya sp.[7]
- Homomya ovalis[101]
- Homomya venulithus[102]
- Homomya centralis[103]
- Homomya librata[71]
- Isognomon sp. [104]
- Leda sp.[7]
- Liastrea hisingeri[105]
- Lima duplicata[104]
- Lima pectinoides[106]
- Limea acuticostata[107]
- Limea katsloesensis[107]
- Liogryphaea sp.[7]
- Liogryphaea arcuata[108]
- Liogryphaea lata[109]
- Lioqryphaea regularis[110]
- Modiola hillana[110]
- Modiola ruuthi[111]
- Modiola cf. tenuissima[112]
- Modiola scalprum[112]
- Myoconcha decorata[113]
- Mytilus cf. lamellosus[114]
- Nucula sp.[7]
- Nucula distinguenda[98]
- Nuculana sp.[7]
- Nuculana zieteni[115]
- Nuculana doris[116]
- Oxytoma sinemurensis[7]
- Oxytoma inaequivalvis[117]
- Oxytoma scanica[117]
- Palaeoneilo sp.[7]
- Palaeoneilo galatea[118]
- Palaeoneilo bornholmiensis[119]
- Palaeoneilo oviformis[115]
- Plagiostoma succincta[106]
- Platymya aquarum[120]
- Pleuromya sp.[7]
- Pleuromya forchhammeri[72]
- Pleuromya corrugata[121]
- Plicatula spinosa[87]
- Plicatula orbiculoides[122]
- Protocardia philippiana[123]
- Protocardia oxynoti[123]
- Protocardia truncata[124]
- Pseudopecten aequivalvis[125]
- Pseudopis sp.[126]
- Rollieria sp.[7]
- Rollieria bronni[127]
- Sphaeriola kurremolinae[123]
- Taeniodon nathorsti[113]
- Tancredia arenacea[126]
- Tancredia securiformis[128]
- Tancredia erdmanni[129]
- Tancredia johnstrupi[129]
- Tancredia lineata[130]
- Terquemia arietis[131]
- Trigonia primaeva[132]
- Trigonia modesta[133]
- Tutcheria cingulata[134]
- Tutcheria cf. rickardsoni[126]
Gastropods
Scaphopods
- Baltodentalium weitschati[137]
- "Dentalium" hexagonale[138]
- Laevidentalium elongatum[138]
- Laevidentalium sp.[139]
- Prodentalium bandeli[140]
- Progadilina spaethi[141]
- Progadilina subtrigonalis[142]
Branchiopoda
- Kuqaia scanicus[143]
Ostracods
- Astacolus denticulina carinata[47]
- Brizalina liassica amalthea[47]
- Citharina inaequistriata[47]
- Cristacythere crassireticulata[55]
- Marginulina spinata spinata[47]
- Nanacythere simplex[47]
- Ogmoconchella danica[55]
- Ogmoconchella mouhersensis[55]
- Ogmoconchella adenticulata[47]
- Ogmoconcha amalthei amalthei[55]
- Pleurifera harpa[55]
Mites
- Hydrozetes nsp.[144]
Ichnofossils
Fish
Apart from a few teeth of the hybodont Hybodus reticulatus, the shark fauna from the Rya Formation is exclusively neoselachian.[146]
Color key
|
Notes Uncertain or tentative taxa are in small text; |
| Genus/Family | Species | Location | Member | Material | Notes/Affinities | Images |
|---|---|---|---|---|---|---|
|
Katslösa, Bed 30 |
Katslösa Member |
|
Interpreted as sagittal otoliths of a Palaeonisciformes Indet.[149] Originally assigned to the extant family Sciaenidae, then from members of Palaeoniscidae and finally some recent research suggest affinities with Pholidophoriformes. Bony Fish Otoliths Similar to ones recovered from the Hasle Formation.[150] |
||
|
Katslösa, Bed 42 |
|
A marine Shark, member of the Acrodontidae. |
|||
|
Road between the hamlets of Rya and Katslosa |
Rydebäck Member |
|
Incertae Sedis Bony Fish Scales |
||
|
|
A Marine Shark of the Family Agaleidae |
||||
|
|
Incertae Sedis fragmentary remains of Chimaeras |
||||
|
|
A marine Shark, member of the Hybodontiformes. Related to Hybodus hauffianus and other genera from the south of Germany. Only non-Neoselachian recovered from the location. |
| |||
|
|
Marine Cow Sharks, Incertae Sedis Inside Hexanchidae. Isolated teeth appear to be similar to Hexanchus arzoeensis. |
%252C_Agaleus_dorsetensis_(D-G)_and_Paraorthacodus_sp._(H-I).jpg.webp) | |||
|
|
Marine Sharks of the Family Palaeospinacidae. One of the earliest records of the genus Paraorthacodus, together with others from France. | ||||
|
Palidiplospinax[153] |
|
|
Marine Sharks of the Family Palaeospinacidae. The complete scale described above most closely resembles scales from Synechodus pinnai. |
_%2526_S._enniskilleni_(H-J).jpg.webp) | ||
|
|
Marine Sharks of the Family Palaeospinacidae. The tooth described may represent a new species. |
||||
|
Sphenodus[154] |
|
|
Marine Sharks of the Family Orthacodontidae. |
 | ||

Plant Remains
Coalified wood occurs as scattered pieces up to 6 centimetres (2.4 in) long and indeterminate belemnites, echinoids, serpulids, ostracods and nodosariid foraminifera were also recorded in the formation.[7] Ammonites where found on layers with plant fossils. Fragments of Dactylioceras were found at a depth of 170 m in the Vilhelmsfalt borehole, together with plant remains and beautifully preserved Pelecypods.[155] The plant material, probably derived from watercourses emptying in the neighborhood, consists mostly of small fragments that seem to have been intimately sedimented with the muddy material that flocculated on meeting the seawater.[155] Other specimen, Arnioceras sp. indet. occurs on what appears to be a transitional environment, probably a back-beach with coalified fragments of plants, some of a large size. Probably due to the shell being washed to land due to a storm.[155]
- Palynology
| Genus | Species | Stratigraphic position | Member | Material | Notes | Images |
|---|---|---|---|---|---|---|
|
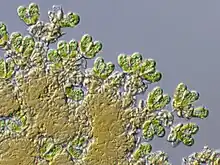
|
Rydebäck Member |
|
Type Genus of the Botryococcaceae inside Trebouxiales. A colonial green microalga related with freshwater and brackish ponds and lakes around the world, where it often can be found in large floating masses. This genus indicates towards the Toarcian section an increasing amount of terrestrial components, as well decreasing salinit in the nearby emerged Skarup Platform.[157] |
 Extant Specimen | |
|
Schizosporis[158] |
|
A Green Algae, member of Zygnemataceae |
||||
|
Tasmanites[26] |
|
A Green Algae, member of Pyramimonadaceae |
||||
|
Campenia[159] |
|
A Green Algae, member of Prasinophyceae |
||||
|
|
A Dinoflajellate, member of the family Nannoceratopsiaceae. It is a genus related with Marine deposits. |
||||
|
Veryhachium[161] |
|
A Dinoflajellate, Dinophyceae familia incertae sedis |
||||
|
|
Affinities with the family Notothyladaceae inside Anthocerotopsida. Hornwort spores. |
).png.webp) Example of extant Notothylas specimens, Foraminisporis come probably from similar genera | |||
|
Neochomotriletes[163] |
| |||||
|
Taurocusporites[25] |
| |||||
|
Polycingulatisporites[164] |
| |||||
|
Staplinisporites[159] |
|
Affinities with the family Encalyptaceae inside Bryopsida. Branching Moss Spores, related with high water-depleting environments |
_2281.JPG.webp) Example of extant Encalypta specimens, Staplinisporites come probably from similar genera | |||
|
Affinities with the family Sphagnaceae inside Sphagnopsida. "Peat moss" spores, related to genera such as Sphagnum that can store water large amount of water. |
 Example of extant Sphagnum specimens, Stereisporites, Sculptisporis and Rogalskaisporites come probably from similar genera | ||||
|
Affinities with the family Lycopodiaceae inside Lycopodiopsida. Lycopod spores, related with herbaceous to arbustive flora common on humid environments |
 Extant Lycopodium specimens. Genera like Lycopodiumsporites probably come from a similar plant | ||||
| ||||||
|
Lycospora[161] |
| |||||
| ||||||
| ||||||
|
Ceratosporites[159] |
|
Affinities with the Selaginellaceae inside Lycopsida. Herbaceous Lycophyte flora, similar to Ferns, ralated with Humid Settings. This Family of Spores are also the most diverse on the Formation. |
 Extant Selaginella, typical example of Selaginellaceae. Genera like Ceratosporites and Densoisporites probably come from a similar or a related Plant | |||
| ||||||
|
Heliosporites[169] |
| |||||
| ||||||
|
Neoraistrickia[170] |
| |||||
|
Uvaesporites[171] |
| |||||
|
Calamospora[172] |
|
Affinities with the Calamitaceae inside Equisetales. Horsetails, herbaceous flora related to high humid environments, flooding tolerant plants. |
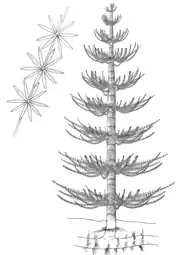 Recosntruction of the Genus Calamites, found associated with Calamospora | |||
|
Incertae Sedis affinities with the Pteridophyta. Uncertain Pteridophyte origin |
|||||
|
Tigrisporites[166] |
| |||||
|
Converrucosisporites[170] |
| |||||
|
Verrucosisporites[161] |
| |||||
|
Cingutriletes[166] |
| |||||
|
Laevigatisporites[161] |
| |||||
|
Convolutispora[164] |
| |||||
| ||||||
|
Simozonotriletes[171] |
| |||||
|
Platyptera[174] |
| |||||
| ||||||
|
Affinities with the family Dennstaedtiaceae inside Polypodiales. Forest Fern Spores |
.jpg.webp) Example of extant Dennstaedtia specimens, Leptolepidites come probably from similar genera | ||||
|
Affinities with the family Botryopteridaceae inside Polypodiales. Forest Fern Spores |
|||||
|
Apiculatisporites[162] |
|
Affinities with the family Zygopteridaceae inside Zygopteridales. |
||||
|
Affinities with the Ophioglossaceae inside Filicales. Fern spores from lower herbaceous flora |
 Example of extant Helminthostachys specimens, Lycopodiacidites come probably from similar genera or maybe a species from the genus | ||||
|
Affinities with the Pteridaceae inside Polypodiopsida. Forest Ferns from humid ground locations |
.jpg.webp) Example of extant Pityrogramma specimens, Contignisporites and Manumia come probably from similar genera or maybe a species from the genus | ||||
|
Auritulinasporites[159] |
|
Affinities with the Gleicheniales inside Polypodiopsida. Fern spores from lower herbaceous flora |
 Example of extant Gleichenia specimens, Gleicheniidites and Auritulinasporites come probably from similar genera or maybe a species from the genus | |||
| ||||||
|
Dictyophyllidites[166] |
|
Affinities with the Dipteridaceae inside Gleicheniales. Fern spores from lower herbaceous flora |
 Example of extant Dipteris specimens, Dictyophyllidites come probably from similar genera or maybe a species from the genus | |||
|
Affinities with the Marattiaceae inside Polypodiopsida. Fern spores from lower herbaceous flora |
.jpg.webp) Example of extant Marattia specimens, Marattisporites come probably from similar genera | ||||
|
Affinities with the Matoniaceae inside Polypodiopsida. Fern spores from lower herbaceous flora |
 Example of extant Matonia specimens, Matonisporites come probably from similar genera | ||||
|
Affinities with the family Osmundaceae inside Polypodiopsida. Near Fluvial currents ferns, reted to the modern Osmunda Regalis. |
_-_Cape_St._Mary's_Ecological_Reserve%252C_Newfoundland_2019-08-10.jpg.webp) Example of extant Osmunda specimens, Osmundacidites and Baculatisporites come probably from similar genera or maybe a species from the genus | ||||
| ||||||
| ||||||
|
Affinities with the family Cyatheaceae inside Cyatheales. Arboreal Fern Spores |
 Example of extant Cyathea, Zebrasporites and Cibotiumspora come probably from similar genera | ||||
| ||||||
| ||||||
| ||||||
|
|
Incertae sedis Affinities with the Gymnospermophyta |
||||
|
Affinities with the families Umkomasiaceae, Peltaspermaceae and Corystospermaceae inside Peltaspermales. Possible seed Fern Pollen |
|||||
|
Affinities with the families Caytoniaceae inside Caytoniales. Possible seed Fern Pollen |
|||||
| ||||||
|
Aratisporites[159] |
| |||||
|
Type Pollen of the Erdtmanithecales, that can be related with the Gnetales. |
|||||
|
Affinities with the family Cycadaceae inside Cycadales. Is among the most abundant flora recovered on the upper section of the formation, and was found to be similar to the pollen of the extant Encephalartos laevifolius. On the Toarcian-Aalenian section, at least six species of Chasmatosporites are recovered, playing more than 50% of the total palynological samples, implicating a clear dominant role of the producer of this Pollen.[184] C. minor is present more on sandy brackish facies, while C. major is found more on brackish-marine facies.[184] |
 Extant Encephalartos laevifolius. Chasmatosporites maybe come from a related plant | ||||
|
Affinities with the Ginkgoaceae inside Ginkgoales. |
|||||
|
Affinities with the Palissyaceae inside Pinopsida. |
|||||
|
Affinities with the Cheirolepidiaceae inside Pinopsida. |
|||||
|
Spheripollenites[178] |
| |||||
|
Affinities with the family Pinaceae inside Pinopsida. Conifer pollen from medium to large arboreal plants |
 Extant Picea. Cerebropollenites and Pinuspollenites maybe come from a related plant | ||||
| ||||||
| ||||||
| ||||||
|
Affinities with the Podocarpaceae inside Pinopsida. Conifer pollen from medium to large arboreal plants |
.jpg.webp) Extant Podocarpus. Podocarpidites and Parvisaccites maybe come from a related plant | ||||
| ||||||
|
Affinities with the family Cupressaceae inside Pinopsida. Pollen that resembles extant genera such as the Genus Actinostrobus and Austrocedrus, probably derived from Dry environments. |
 Extant Austrocedrus. Exesipollenites and Perinopollenites maybe come from a related plant | ||||
| ||||||
|
Affinities with the family Araucariaceae inside Pinales. Conifer Pollen from medium to large Arboreal Plants |
 Extant Araucaria. Araucariacites and Callialasporites maybe come from a related plant | ||||
| ||||||
- Plant Macrofossils
| Genus | Species | Stratigraphic position | Member | Material | Notes | Images |
|---|---|---|---|---|---|---|
|
|
Katslösa Member |
|
A fern of the family Polypodiidae inside Polypodiopsida. Similar to Sphenopteris species of Permian rocks of the same region |
.jpg.webp) Specimen | |
|
|
A member of the family Ginkgoaceae inside Ginkgoales. The shape and the clearly visible bifurcation of the veins agrees with the referral to the genus Ginkgoites |
||||
|
Marchantiolites[190] |
|
|
A member of the family Marchantiaceae inside Hepatophyta. Suggested initially to be an angiosperm leaf. Like extant Marchantia polymorpha, mostly blackish stripes appear along the midrib of the thallus. |
 Extant Marchantia polymorpha | ||
See also
- List of fossiliferous stratigraphic units in Sweden
- Kristianstad Basin
- Toarcian formations
- Djupadal Formation, Central Skane
- Marne di Monte Serrone, Italy
- Calcare di Sogno, Italy
- Mizur Formation, North Caucasus
- Sachrang Formation, Austria
- Saubach Formation, Austria
- Posidonia Shale, Lagerstätte in Germany
- Ciechocinek Formation, Germany and Poland
- Krempachy Marl Formation, Poland and Slovakia
- Lava Formation, Lithuania
References
- ↑ Ahlberg et al., 2003, p.528
- 1 2 Ahlberg et al., 2003, p.530
- 1 2 3 4 Ahlberg et al., 2003, p.533
- ↑ Erlström et al., 2018, p.129
- ↑ Hinz-Schallreuter & Schallreuter, 2009, p.2
- 1 2 3 4 Andersson & Hybertsen, 2010-8-17
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 Frandsen & Surlyk, 2003, p.547
- ↑ Frandsen & Surlyk, 2003, p.550
- 1 2 3 4 5 Ahlberg et al., 2003, p.534
- ↑ Greiff, 2019-8
- ↑ Nielsen et al., 2003, p.588
- ↑ Nielsen et al., 2003, p.590
- ↑ Frandsen & Surlyk, 2003, p.543
- ↑ Erlström et al., 2018, p.138
- ↑ Bryan & Ferrari, 2013, p.1058
- ↑ Bergelin, 2009
- ↑ Augustsson, 2009
- 1 2 Ahlberg et al., 2003, p.539
- ↑ Harries & Little, 1999
- ↑ Erlström et al., 2018, p.139
- ↑ Sjöberg, 2018, p.90
- ↑ Bou Daher (2012)-p10
- ↑ Bou Daher (2012)-p7
- ↑ Bou Daher (2012)-p37
- 1 2 3 4 5 6 7 8 9 10 11 12 13 Guy-Ohlson, 1986-p3
- 1 2 3 4 5 6 7 8 Guy-Ohlson, 1990-p218
- ↑ Bou Daher (2012)-p14
- ↑ Bou Daher (2012)-p15
- 1 2 Seidenkrantz & Koppelhus (1993)-p201
- 1 2 3 Seidenkrantz & Koppelhus (1993)-p210
- 1 2 Nagy, J. (2003)-p28
- 1 2 Ahlberg, Olsson & Šimkevičius, 2003-p15
- ↑ Ahlberg, Olsson & Šimkevičius, 2003-p17
- ↑ Ahlberg, Olsson & Šimkevičius, 2003-p19
- ↑ Ahlberg, Olsson & Šimkevičius, 2003-p21
- ↑ Dera, Prunier, Smith, Haggart, Popov, Guzhov, Rogovf, Delsate, Detlev,Pucéatj, Charbonnierk & Germain (2015)-p1610
- 1 2 3 4 Hunter, A.W. & Rees, J.(2010)-p10-24
- ↑ Augustsson (2001)-p28
- ↑ Bölau & Kockel-Brosius (1965)-p55
- ↑ Dera, Prunier, Smith, Haggart, Popov, Guzhov, Rogovf, Delsate, Detlev,Pucéatj, Charbonnierk & Germain (2015)-p1611
- ↑ Guy-Ohlson, 1988-p11
- ↑ Rya Formation at Fossilworks.org
- ↑ Weitschat & Gründel (2002)-p39
- 1 2 3 Troedsson (1951)-p146-147
- 1 2 Troedsson (1951)-p15
- ↑ Reyment (1959)-p118
- 1 2 3 4 5 6 7 8 9 10 11 Sivhed, U. 1984-p19
- ↑ Doguzhaeva, Mutvei & Weitschat (2003)-p83
- ↑ Reyment (1959)-p129
- ↑ Reyment (1959)-p130
- 1 2 3 4 5 6 Frandsen & Surlyk, 2003, p.545
- ↑ Reyment (1959)-p110
- ↑ Reyment (1959)-p134
- ↑ Reyment (1959)-p142
- 1 2 3 4 5 6 7 8 9 Sivhed, U. (1984)- p.251
- ↑ Lehmann (1961)-p42-43
- ↑ Reyment (1959)-p132
- ↑ Reyment (1959)-p112
- ↑ Reyment (1959)-p136
- ↑ Reyment (1959)-p114
- ↑ Doguzhaeva, Mutvei & Weitschat (2003)-p86
- ↑ Reyment (1959)-p139
- ↑ Reyment (1959)-p137
- ↑ Doguzhaeva, Mutvei & Weitschat (2003)-p80
- ↑ Doguzhaeva, Mutvei & Weitschat (2003)-p81
- 1 2 3 4 5 Troedsson (1951)-p243-245
- ↑ Doguzhaeva, Mutvei & Weitschat (2003)-p79
- 1 2 3 4 Troedsson (1951)-p148-150
- ↑ Troedsson (1951)-p224
- 1 2 Troedsson (1951)-p185
- 1 2 Troedsson (1951)-p192
- 1 2 Troedsson (1951)-p193
- ↑ Troedsson (1951)-p167
- 1 2 Troedsson (1951)-p168
- ↑ Troedsson (1951)-p169
- ↑ Troedsson (1951)-p170
- 1 2 Troedsson (1951)-p171
- ↑ Troedsson (1951)-p159
- ↑ Troedsson (1951)-p160
- ↑ Troedsson (1951)-p161
- ↑ Troedsson (1951)-p162
- ↑ Troedsson (1951)-p163
- ↑ Troedsson (1951)-p211
- ↑ Troedsson (1951)-p212
- 1 2 Troedsson (1951)-p213
- ↑ Troedsson (1951)-p214
- 1 2 Troedsson (1951)-p220
- 1 2 Troedsson (1951)-p216
- ↑ Troedsson (1951)-p217
- ↑ Troedsson (1951)-p218
- 1 2 Troedsson (1951)-p181
- ↑ Troedsson (1951)-p184
- ↑ Troedsson (1951)-p203
- ↑ Troedsson (1951)-p204
- ↑ Troedsson (1951)-p205
- ↑ Troedsson (1951)-p206
- ↑ Troedsson (1951)-p195
- 1 2 Troedsson (1951)-p154
- ↑ Troedsson (1951)-p157
- ↑ Troedsson (1951)-p158
- ↑ Troedsson (1951)-p188
- ↑ Troedsson (1951)-p189
- ↑ Troedsson (1951)-p191
- 1 2 Troedsson (1951)-p207
- ↑ Troedsson (1951)-p225
- 1 2 Troedsson (1951)-p208
- 1 2 Troedsson (1951)-p210
- ↑ Troedsson (1951)-p226
- ↑ Troedsson (1951)-p227
- 1 2 Troedsson (1951)-p228
- ↑ Troedsson (1951)-p231
- 1 2 Troedsson (1951)-p232
- 1 2 Troedsson (1951)-p234
- ↑ Troedsson (1951)-p233
- 1 2 Troedsson (1951)-p151
- ↑ Troedsson (1951)-p152
- 1 2 Troedsson (1951)-p200
- ↑ Troedsson (1951)-p149
- ↑ Troedsson (1951)-p150
- ↑ Troedsson (1951)-p187
- ↑ Troedsson (1951)-p194
- ↑ Troedsson (1951)-p221
- 1 2 3 Troedsson (1951)-p178
- ↑ Troedsson (1951)-p179
- ↑ Troedsson (1951)-p219
- 1 2 3 Troedsson (1951)-p173
- ↑ Troedsson (1951)-p153
- ↑ Troedsson (1951)-p175
- 1 2 Troedsson (1951)-p176
- ↑ Troedsson (1951)-p177
- ↑ Troedsson (1951)-p223
- ↑ Troedsson (1951)-p165
- ↑ Troedsson (1951)-p166
- ↑ Troedsson (1951)-p172
- 1 2 3 4 5 6 7 Troedsson (1951)-p238-240
- ↑ Weitschat & Gründel (2002)-p40
- ↑ Engeser & Riedel (1992)-p49-50
- 1 2 Troedsson (1951)-p230-245
- ↑ Engeser & Riedel (1992)-p46
- ↑ Engeser & Riedel (1992)-p43
- ↑ Engeser & Riedel (1992)-p47
- ↑ Engeser & Riedel (1992)-p48
- ↑ Peng, Slater, McLoughlin & Vajda, (2023)-p4-20
- ↑ Sivhed & Wallwork (1978)-p65-66
- 1 2 3 4 5 6 Frandsen & Surlyk, 2003, p.546
- ↑ Rees, 2000, p.411
- 1 2 Troedsson (1951)-p246
- ↑ Schwarzhans (2018)-p79
- ↑ Schwarzhans (2018)-p112
- ↑ Schwarzhans (2018)-p83
- 1 2 3 4 5 6 Rees, 2000, p.412
- ↑ Rees, 2000, p.416
- ↑ Rees, 2000, p.417-18
- ↑ Rees, 2000, p.415
- 1 2 3 Reyment (1959)-p152
- ↑ Guy-Ohlson, 1996-p250
- ↑ Guy-Ohlson, 1996-p252
- ↑ Guy-Ohlson, 1990-p217
- 1 2 3 4 5 6 7 8 9 10 11 Guy-Ohlson, 1986-p2
- 1 2 3 4 5 6 7 Guy-Ohlson, 1990-p220
- 1 2 3 4 5 6 7 8 Guy-Ohlson, 1990-p222
- 1 2 3 4 5 6 Guy-Ohlson, 1990-p219
- 1 2 3 4 5 6 7 Guy-Ohlson, 1986-p4
- 1 2 3 4 Guy-Ohlson, 1990-p221
- 1 2 3 4 Guy-Ohlson, 1986-p5
- 1 2 3 4 5 6 7 8 9 10 Guy-Ohlson, 1990-p223
- 1 2 3 Guy-Ohlson, 1990-p224
- 1 2 3 4 5 Guy-Ohlson, 1986-p6
- 1 2 3 4 5 Guy-Ohlson, 1990-p225
- 1 2 3 Guy-Ohlson, 1986-p7
- 1 2 3 Guy-Ohlson, 1986-p10
- 1 2 Guy-Ohlson, 1986-p9
- ↑ Guy-Ohlson, 1988-p807
- 1 2 3 Guy-Ohlson, 1986-p11
- ↑ Guy-Ohlson, 1988-p808
- ↑ Guy-Ohlson, 1986-p8
- 1 2 3 4 5 6 7 Guy-Ohlson, 1990-p226
- 1 2 Guy-Ohlson, 1986-p12
- ↑ Guy-Ohlson, 1986-p16
- 1 2 Guy-Ohlson, 1990-p227
- 1 2 3 4 5 6 Guy-Ohlson, 1986-p15
- ↑ Guy-Ohlson, 1988-p809
- ↑ Guy-Ohlson, 1988-p810
- 1 2 Guy-Ohlson, 1988-p6-12
- ↑ Guy-Ohlson, 1988-p812
- ↑ Guy-Ohlson, 1988-p813
- ↑ Guy-Ohlson, 1988-p815
- ↑ Roselt (1968)-p65
- ↑ Roselt (1968)-p66
- ↑ Roselt (1968)-p68
Bibliography
- Peng, J.; Slater, S. M.; McLoughlin, S.; Vajda, V. (2023). "New species of Kuqaia from the Lower Jurassic of Sweden indicates a possible water flea (Crustacea: Branchiopoda) affinity". PLOS ONE. 18 (6): 4–20. doi:10.1371/journal.pone.0282247. PMC 10246822. PMID 37285340.
- Greiff, Johannes (2019). "Mesozoiska konglomerat och Skånes tektoniska utveckling". Examensarbeten I Geologi Vid Lunds Universitet. 18 (1): 8–17. Retrieved 20 January 2021.
- Erlström, M.; L.O. Boldreel; S. Lindström; L. Kristensen; A. Mathiesen; M.S. Andersen; E. Kamla, and L.H. Nielsen. 2018. Stratigraphy and geothermal assessment of Mesozoic sandstone reservoirs in the Øresund Basin – exemplified by well data and seismic profiles. Bulletin of the Geological Society of Denmark 66. 123–149. Accessed 2020-07-09. ISSN 2245-7070
- Schwarzhans, W. (2018). "A review of Jurassic and Early Cretaceous otoliths and the development of early morphological diversity in otoliths". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 287 (1): 75–121. doi:10.1127/njgpa/2018/0707. Retrieved 13 August 2021.
- Sjöberg, Björn. 2018. Strategic Environmental Assessment of the Marine Spatial Plan proposal for Skagerrak and Kattegat, 1–129. Swedish Agency for Marine and Water Management. Accessed 2020-07-09.
- Sachs, S.; Hornung, J. J.; Lierl, H. J; Kear, B. P. (2016). "Plesiosaurian fossils from Baltic glacial erratics: evidence of Early Jurassic marine amniotes from the southwestern margin of Fennoscandia". Geological Society, London, Special Publications. 434 (1): 149–163. Bibcode:2016GSLSP.434..149S. doi:10.1144/SP434.14. S2CID 130195351.
- Dera, Guillaume; Prunier, Jonathan; Smith, Paul L.; Haggart, James W.; Popov, Evgeny; Guzhov, Alexander; Rogovf, Mikhail; Delsateg, Dominique; Thiesh, Detlev; Cunyi, Gilles; Pucéatj, Emmanuelle; Charbonnierk, Guillaume; Bayonl, Germain (June 2015). "Nd isotope constraints on ocean circulation, paleoclimate, and continental drainage during the Jurassic breakup" (PDF). Gondwana Research. 2 (4): 1599–1615. doi:10.1016/j.gr.2014.02.006. Retrieved 11 August 2021.
- Bryan, S. E., and L. Ferrari. 2013. Large igneous provinces and silicic large igneous provinces: Progress in our understanding over the last 25 years. Geological Society of America Bulletin 125(7-8). 1053–1078. Accessed 2020-07-09. doi:10.1130/B30820.1
- Bou Daher, Samer (2012). "Lithofacies analysis and heterogeneity study of the subsurface Rhaetian– Pliensbachian sequence in SW Skåne and Denmark". Dissertations in Geology at Lund University (1): 1–55. Retrieved 9 May 2021.
- Andersson, Josefin; Hybertsen, Frida (2010). "Geologi i Helsingborgs kommun: en geoturistkarta med beskrivning". Geologi I Helsingborgs Kommun – en Geoturistkarta Med Beskrivning. 15 (272): 7–8. Retrieved 20 January 2021.
- Bergelin, Ingemar. 2009. Jurassic volcanism in Skåne, southern Sweden, and its relation to coeval regional and global events. GFF 131(1–2). 165–175. Accessed 2020-07-09. doi:10.1080/11035890902851278
- Hunter, A.W., and J. Rees. 2010. A new echinoderm faunule from the Lower Jurassic (Pliensbachian) of southern Sweden. Bulletin of the Geological Society of Denmark 58. 66-73. . ISSN 0011-6297 E-ISSN 2245-7070
- Hinz-Schallreuter, Ingelore, and Roger Schallreuter. 2009. Geschiebe-Oolithe und –Onkolithe III - Mesozoische Oolithe - Oolites and Onkolites as Geschiebes (glacial erratic boulders) III - Mesozoic Oolites. Geschiebekunde aktuell 25. 1–10. Accessed 2020-07-09.
- Doguzhaeva, L. A.; Mutvei, H.; Weitschat, W. (2003). "The pro-ostracum and primordial rostrum at early ontogeny of Lower Jurassic belemnites from north-western Germany". Berliner Paläobiologische Abhandlungen. 3 (1): 79. Retrieved 16 July 2021.
- Ahlberg, A.; Olsson, I.; Šimkevičius, P. (2003). "Triassic–Jurassic weathering and clay mineral dispersal in basement areas and sedimentary basins of southern Sweden". Sedimentary Geology. 161 (1–2): 15. Bibcode:2003SedG..161...15A. doi:10.1016/S0037-0738(02)00381-0.
- Nagy, J (2003). "New foraminiferal taxa and revised biostratigraphy of Jurassic marginal marine deposits on Anholt, Denmark". Micropaleontology. 49 (1): 27–46. doi:10.2113/49.1.27.
- Ahlberg, Anders; Ulf Sivhed, and Mikael Erlström. 2003. The Jurassic of Skåne, southern Sweden. Geological Survey of Denmark and Greenland Bulletin 1. 527–541. Accessed 2020-07-09.
- Frandsen, Nils, and Finn Surlyk. 2003. An offshore transgressive-regressive mudstone-dominated succession from the Sinemurian of Skåne, Sweden. Geological Survey of Denmark and Greenland Bulletin 1. 585–609. Accessed 2020-07-09.
- Weitschat, W.; Gründel, J. (2002). "Eine agglutinierte Wurmröhre (Polychaeta, Sedentaria, Terebellomorpha) aus einem Geschiebe des Mittel-Lias (Domerium, spinatum Zone) Norddeutschlands". Mitt. Geol-Paläont Inst Univ Hamburg. 86 (1): 37.
- Augustsson, Carita. 2001. Lapilli tuff as evidence of Early Jurassic Strombolian-type volcanism in Scania, southern Sweden. GFF 123(1). 23–28. Accessed 2020-07-09. doi:10.1080/11035890101231023
- Rees, Jan. 2000. A new Pliensbachian (Early Jurassic) neoselachian shark fauna from southern Sweden. Acta Palaeontologica Polonica 45. 407–424. Accessed 2020-07-09.
- Harries, Peter J., and Crispin T.S. Little. 1999. The early Toarcian (Early Jurassic) and the Cenomanian–Turonian (Late Cretaceous) mass extinctions: similarities and contrasts. Palaeogeography, Palaeoclimatology, Palaeoecology 154(1–2). 39–66. . doi:10.1016/S0031-0182(99)00086-3 Bibcode:1999PPP...154...39H
- Guy-Ohlson, D. (1996). "The Significance of Specific Microalgae in the Palaeoenvironmental Interpretation and Biostratigraphical Correlation of the Jurassic in Sweden". In Geo Research Forum. 13 (3): 249–254.
- Seidenkrantz, M. S.; Koppelhus, E. B. (1993). "Biostratigraphy and palaeoenvironmental analysis of a Lower to Middle Jurassic succession on Anholt, Denmark". Journal of Micropalaeontology. 12 (2): 201–218. Bibcode:1993JMicP..12..201S. doi:10.1144/jm.12.2.201.
- Guy-Ohlson, D.. 1990. Pliensbachian palynology of the Karindal bore no. 1, north-west Scania, Sweden.. Review of Palaeobotany and Palynology 65(4). 217-228. .
- Engeser, T; Riedel, F (1992). "Scaphopods from middle Liassic erratic boulders of northern Germany: with a review on Liassic Scaphopoda". Scripta Geologica. 99 (1): 35.
- Guy-Ohlson, D. (1988). "Toarcian palynostratigraphical correlations within and between different biogeographical Provinces". International Symposium on Jurassic Stratigraphy. 1 (1): 807–820.
- Guy-Ohlson, D.. 1988. The use of dispersed palynomorphs referable to the form genus Chasmatosporites (Nilsson) Pocock and Jansonius, in Jurassic biostratigraphy. Congreso Argentino de Paleontologia y Bioestratigrafia 3(1–2). 5- 13. Accessed 09 April 2021.
- Guy-Ohlson, D.. 1985. Jurassic Palynology of the Vilhelmsfält Bore No. 1, Scania, Sweden: Toarcian-Aalenian. Section of Palaeobotany, Swedish Museum of Natural History 86(2). 1-18. .
- Sivhed, Ulf (1984). "Litho-and biostratigraphy of the Upper Triassic-Middle Jurassic in Scania, southern Sweden " (PDF). Sveriges geologiska undersökning. 31 (12): 31–50. Retrieved 11 February 2021.
- Sivhed, Ulf (1981). "Stratigraphy of the Gantofta-Katslösa area in Scania, Sweden". Geologiska Föreningen i Stockholm Förhandlingar. 103 (2): 249–252. doi:10.1080/11035898109454521.
- Sivhed, U.; Wallwork, J. A. (1978). "An early Jurassic oribatid mite from southern Sweden". Geologiska Föreningen i Stockholm Förhandlingar. 100 (1): 65–70. doi:10.1080/11035897809448562. Retrieved 24 December 2021.
- Lehmann, Ulrich (1971). "Faziesanalyse der Ahrensburger Liasknollen auf Grund ihrer Wirbeltierreste". Mitteilungen aus dem Geologischen Institut der Technischen Universität Hannover. 10 (1): 21.
- Roselt, G. (1968). "Jurassische Pflanzenreste aus einem Geschiebe-Geröll vom Ostseestrand der Mecklenburger Bucht". Botanical Journal of the Linnean Society. 61 (1): 65–74. doi:10.1111/j.1095-8339.1968.tb00103.x.
- Bölau, E.; Kockel-Brosius, M. (1965). "Der tertiäre Vulkanismus in Zentralschonen, Südschweden: Mikropaläontologische Untersuchungen an Bohrproben aus Schonen" (PDF). Gleerup (1): 1–55. Retrieved 30 July 2021.
- Lehmann, Ulrich (1961). "Ammoniten mit Kieferapparat und Radula aus Lias-Geschieben". Paläontologische Zeitschrift. 41 (1): 38. doi:10.1007/BF02998547. S2CID 128711331.
- Reyment, R. A. (1959). "On liassic ammonites from Skåne, southern Sweden" (PDF). Almqvist & Wiksell. 1 (5): 105. Retrieved 13 July 2021.
- Troedsson, Gustaf (1951). On the Höganäs series of Sweden (RhaetoLias) (PDF) (1 ed.). Sweden: CWK Gleerup. p. 119. Retrieved 19 January 2021.
External links
![]() Media related to Rya Formation at Wikimedia Commons
Media related to Rya Formation at Wikimedia Commons
| Ph |
| ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|