| Reverse gyrase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
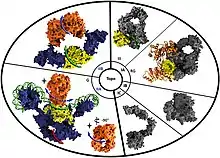 Molecular depictions of the different type I and type II topoisomerase classes. Reverse gyrase is indicated by the 'RG' tag under the type IA topoisomerase section of the diagram. | |||||||||
| Identifiers | |||||||||
| EC no. | 5.6.2.2 | ||||||||
| CAS no. | 143180-75-0 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| |||||||||
Reverse gyrase is a type I topoisomerase that introduces positive supercoils into DNA,[1] contrary to the typical negative supercoils introduced by the type II topoisomerase DNA gyrase. These positive supercoils can be introduced to DNA that is either negatively supercoiled or fully relaxed.[2] Where DNA gyrase forms a tetramer and is capable of cleaving a double-stranded region of DNA, reverse gyrase can only cleave single stranded DNA.[3][4] More specifically, reverse gyrase is a member of the type IA topoisomerase class; along with the ability to relax negatively or positively supercoiled DNA[5] (which does not require ATP[6][7]), type IA enzymes also tend to have RNA-topoisomerase activities. These RNA topoisomerases help keep longer RNA strands from becoming tangled in what are referred to as "pseudoknots." Due to their ability to interact with RNA, it is thought that this is one of the most ancient class of enzymes found to date.[4]
Reverse gyrase is an ATP-dependent topoisomerase[8] in terms of its positive supercoiling activity, however, reverse gyrase can also relax DNA strands without introducing positive supercoils through interaction with ADP.[9] The structure of the enzyme includes both a helicase domain, which is responsible for separating nucleic acids, and a topoisomerase domain, which is responsible for the actual introduction of coils into DNA. However, mechanistic studies have shown that these two domains tend to exhibit weak activities separately and can only perform efficient DNA positive supercoiling activity when working in tandem.[10][11] Other studies have also shown that reverse gyrase enzymes tend to favorably attack regions of single-stranded DNA versus double-stranded DNA, which suggests that this enzyme's critical biological function is to ensure the constant renaturation of melted DNA strands, especially in organisms that grow at high temperatures.[8]
This enzyme has been extensively characterized across several Archaea, with Sulfolobus acidocaldarius reverse gyrase being one of the first to be characterized.[12] Additionally, it has been found that all thermophilic bacteria and archaea contain at least one reverse gyrase enzyme. Some organisms, such as members of the Crenarchaeota phylum, even have two reverse gyrase enzymes: TopR1, which tends to be active in increased temperatures, and TopR2, which shows activity in both low and high temperatures.[4] Other exceptional organisms include Nanoarchaeum equitans, whose reverse gyrase enzyme tends to naturally exist as two separate peptides versus the typical monomeric polypeptide with a topoisomerase IA domain and a helicase domain.[4][11]
Classification
As seen in the information box above, reverse gyrase is designated under the EC number 5.6.2.2. The first number of this code (5) designates the enzymes identity as an isomerase.[13] While the enzyme itself does have both a topoisomerase and helicase-like domain, as a gyrase, it is primarily classified under the topoisomerase umbrella. Furthermore, the 5.6 number designates this molecule as an isomerase that is capable of changing conformation in cellular molecules.[14] 5.6.2 designates the enzyme further as being capable of altering nucleic acid, or DNA, conformations.[15] Lastly, the full designation of 5.6.2.2 characterizes this enzyme as an ATP-dependent DNA topoisomerase.[16]
Structure
The crystal structure of reverse gyrase has been characterized fully, and a crystal structure has been produced based on the enzyme found in Thermotoga maritima.[12]
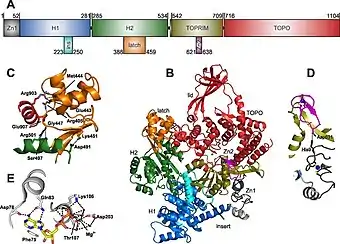
Active Site
Reverse gyrases have helicase and topoisomerase domains. The active site, where nucleotides are bound by the enzyme, is characterized by Asp78, Phe75, Gln83, Lys106, Asp203, and Thr107 residues.[12] It is hypothesized that the H1 and H2 subdomains also contain nucleotide-binding abilities, and the DNA strand is able to be grabbed by these subdomains and are subsequently fed up through the topoisomerase domain of the enzyme to complete positive supercoiling.[12]
Latch Domain
The latch domain appears to be variable across species, with domain size ranging from as small as 10 amino acids to as large as 120 amino acids.[12] The latch is thought to function as a control mechanism to prevent the topoisomerase domain from creating negative supercoils and relaxing the DNA, and instead allows the enzyme to create positive supercoils in an ATP-dependent manner during the strand passage step of the helicase domain.[5][17]
Zinc Fingers
The reverse gyrase enzyme contains a zinc finger domain, where two zinc ions help to coordinate enzymatic function. The first zinc ion is kept in place by interactions with four cysteine residues.[12] The second zinc ion is not always found in reverse gyrase enzymes. However, when present, both ions are found near the binding site for nucleic acids. It is thought that these zinc fingers play a role in initial binding of DNA and strand passage, but their exact mechanisms of action appear to vary between organisms.[12]
Thermophile significance
Organisms that live under standard temperature and pressure conditions, or mesophiles (living in temperature ranges between 20 °C and 40 °C), tend to have negative supercoiling in their DNA strands. This helps to condense the genetic material so that it fits within the host cells (or in the case of eukaryotes, within the cell's nuclear region). Negative supercoiling, also referred to as underwinding, results in the counterclockwise twisting of the DNA strand. Negative supercoiling leaves the DNA strands available for various cellular processes, like genome replication and transcription, as DNA typically needs to be underwound in order to be denatured and accessed by the proper enzymes.[2]
On the other hand, thermophiles (organisms that can live in temperatures ranging from 40 °C up to as high as 122 °C[18]) are thought to maintain several positive supercoils in their DNA in order to assist with maintaining structural integrity of the DNA under the denaturing capabilities of these high temperatures. Positive supercoiling, which is referred to as overwinding, results in the clockwise twisting of the strand. As previously discovered, one of the biggest benefits to maintaining positive supercoils in DNA strands is preventing separation of the strands in high temperatures.[2]
While positive supercoiling is certainly more common in thermophiles, positive supercoiling has been found in mesophilic organisms. For example, telomeres and condensins can both utilize positive supercoiling as a means for contributing to chromosomal structure.[19] Furthermore, the reverse gyrase enzyme is not exclusive to thermophiles. Some reverse gyrase enzymes even function outside of thermophilic temperature ranges, suggesting that there may be some organisms at mesophilic temperatures that utilize this enzyme.[4]
Supercoiling mechanism
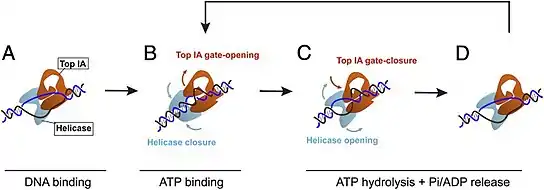
It is suspected that the helicase and topoisomerase domains of the reverse gyrase enzyme work together to promote positive supercoiling in DNA. However, the exact mechanisms of action appear to differ between organisms. For example, Sulfolobus solfataricus and Thermotoga maritima experience opposite phenomena in terms of helicase activity: Sulfolobus solfataricus helicase's ability to hydrolyze ATP appears to be activated by the topoisomerase, whereas Thermotoga maritima ATP hydrolysis ability via the helicase appears to be reduced by the topoisomerase domain.[12]
When bound to the DNA, reverse gyrase induces a change in structure via a left-handed wrapping, which more or less functions as an unwinding.[20] Specifically, the reverse gyrase found in S. solfataricus (a Crenarchaeota TopR2 reverse gyrase) initiates an unwinding of approximately 20 base pairs upon binding to a DNA structure.[4] Upon initial binding to the DNA, the helicase domain is in an open conformation, while the topoisomerase IA domain is in a closed conformation.[5] After the binding of ATP to the reverse gyrase structure, the helicase domain closes, and the topoisomerase IA domain opens. This triggers a rewinding of 10 of the 20 base pairs in the unwound bubble, and the topoisomerase IA domain can introduce positive supercoiling during strand passage.[5] As the strand passage occurs, reverse gyrase's topoisomerase IA domain is able to increase the linking number (how many times a strand of DNA is wrapped around the other strand) of the DNA strand as they are renatured.[9] Following ATP hydrolysis-induced rewinding, the reverse gyrase enzyme domains return to their original state (open helicase domain and closed topoisomerase IA domain) and the reverse gyrase is released, ready to bind to a new region of DNA and repeat the process.[4][5][9]
Regardless of the differences in interactions between the topoisomerase and helicase domains, in general, reverse gyrase enzymes all undergo conformational changes when nucleotides are bound to the active site.[12]
References
- ↑ Kikuchi A, Asai K (21 June 1984). "Reverse gyrase--a topoisomerase which introduces positive superhelical turns into DNA". Nature. 309 (5970): 677–681. Bibcode:1984Natur.309..677K. doi:10.1038/309677a0. PMID 6328327. S2CID 4242694.
- 1 2 3 Fogg JM, Catanese DJ, Randall GL, Swick MC, Zechiedrich L (2009). Benham CJ, Harvey S, Olson WK, Sumners WL, Swigon D (eds.). "Differences Between Positively and Negatively Supercoiled DNA that Topoisomerases May Distinguish". The IMA Volumes in Mathematics and Its Applications. 150: 73–121. doi:10.1007/978-1-4419-0670-0_5. ISBN 978-1-4419-0669-4.
- ↑ Jaxel C, Nadal M, Mirambeau G, Forterre P, Takahashi M, Duguet M (October 1989). "Reverse gyrase binding to DNA alters the double helix structure and produces single-strand cleavage in the absence of ATP". The EMBO Journal. 8 (10): 3135–3139. doi:10.1002/j.1460-2075.1989.tb08466.x. PMC 401394. PMID 2555155.
- 1 2 3 4 5 6 7 Garnier F, Couturier M, Débat H, Nadal M (25 May 2021). "Archaea: A Gold Mine for Topoisomerase Diversity". Frontiers in Microbiology. 12: 661411. doi:10.3389/fmicb.2021.661411. PMC 8185306. PMID 34113328.
- 1 2 3 4 5 Yang X, Garnier F, Débat H, Strick TR, Nadal M (May 2020). "Direct observation of helicase-topoisomerase coupling within reverse gyrase". Proceedings of the National Academy of Sciences of the United States of America. 117 (20): 10856–10864. Bibcode:2020PNAS..11710856Y. doi:10.1073/pnas.1921848117. PMC 7245102. PMID 32371489.
- ↑ "EC 5.6.2.1". IUBMB Enzyme Nomenclature. International Union of Biochemistry and Molecular Biology.
- ↑ Gellert M (1981). "DNA topoisomerases". Annual Review of Biochemistry. 50: 879–910. doi:10.1146/annurev.bi.50.070181.004311. PMID 6267993.
- 1 2 Shibata T, Nakasu S, Yasui K, Kikuchi A (August 1987). "Intrinsic DNA-dependent ATPase activity of reverse gyrase". The Journal of Biological Chemistry. 262 (22): 10419–10421. doi:10.1016/S0021-9258(18)60974-3. PMID 3038879.
- 1 2 3 Hsieh TS, Plank JL (March 2006). "Reverse gyrase functions as a DNA renaturase: annealing of complementary single-stranded circles and positive supercoiling of a bubble substrate". The Journal of Biological Chemistry. 281 (9): 5640–5647. doi:10.1074/jbc.M513252200. PMID 16407212.
- ↑ Déclais AC, Marsault J, Confalonieri F, de La Tour CB, Duguet M (June 2000). "Reverse gyrase, the two domains intimately cooperate to promote positive supercoiling". The Journal of Biological Chemistry. 275 (26): 19498–19504. doi:10.1074/jbc.M910091199. PMID 10748189.
- 1 2 Capp C, Qian Y, Sage H, Huber H, Hsieh TS (December 2010). "Separate and combined biochemical activities of the subunits of a naturally split reverse gyrase". The Journal of Biological Chemistry. 285 (51): 39637–39645. doi:10.1074/jbc.M110.173989. PMC 3000944. PMID 20929866.
- 1 2 3 4 5 6 7 8 9 Lulchev P, Klostermeier D (July 2014). "Reverse gyrase--recent advances and current mechanistic understanding of positive DNA supercoiling". Nucleic Acids Research. 42 (13): 8200–8213. doi:10.1093/nar/gku589. PMC 4117796. PMID 25013168.
- ↑ "EC 5. Isomerases". IUBMB Enzyme Nomenclature. International Union of Biochemistry and Molecular Biology.
- ↑ "EC 5.6". IUBMB Enzyme Nomenclature. International Union of Biochemistry and Molecular Biology.
- ↑ "EC 5.6.2 Enzymes altering nucleic acid conformation". IUBMB Enzyme Nomenclature. International Union of Biochemistry and Molecular Biology.
- ↑ "5.6.2.2". IUBMB Enzyme Nomenclature. International Union of Biochemistry and Molecular Biology.
- ↑ Rodríguez AC, Stock D (February 2002). "Crystal structure of reverse gyrase: insights into the positive supercoiling of DNA". The EMBO Journal. 21 (3): 418–426. doi:10.1093/emboj/21.3.418. PMC 125824. PMID 11823434.
- ↑ Takai K, Nakamura K, Toki T, Tsunogai U, Miyazaki M, Miyazaki J, et al. (August 2008). "Cell proliferation at 122 degrees C and isotopically heavy CH4 production by a hyperthermophilic methanogen under high-pressure cultivation". Proceedings of the National Academy of Sciences of the United States of America. 105 (31): 10949–10954. doi:10.1073/pnas.0712334105. PMC 2490668. PMID 18664583.
- ↑ Valenti A, Perugino G, Rossi M, Ciaramella M (January 2011). "Positive supercoiling in thermophiles and mesophiles: of the good and evil". Biochemical Society Transactions. 39 (1): 58–63. doi:10.1042/BST0390058. PMID 21265747.
- ↑ Confalonieri F, Elie C, Nadal M, de La Tour C, Forterre P, Duguet M (May 1993). "Reverse gyrase: a helicase-like domain and a type I topoisomerase in the same polypeptide". Proceedings of the National Academy of Sciences of the United States of America. 90 (10): 4753–4757. Bibcode:1993PNAS...90.4753C. doi:10.1073/pnas.90.10.4753. PMC 46591. PMID 8389456.