RNA-directed DNA methylation (RdDM) is a biological process in which non-coding RNA molecules direct the addition of DNA methylation to specific DNA sequences. The RdDM pathway is unique to plants, although other mechanisms of RNA-directed chromatin modification have also been described in fungi and animals. To date, the RdDM pathway is best characterized within angiosperms (flowering plants), and particularly within the model plant Arabidopsis thaliana. However, conserved RdDM pathway components and associated small RNAs (sRNAs) have also been found in other groups of plants, such as gymnosperms and ferns. The RdDM pathway closely resembles other sRNA pathways, particularly the highly conserved RNAi pathway found in fungi, plants, and animals. Both the RdDM and RNAi pathways produce sRNAs and involve conserved Argonaute, Dicer and RNA-dependent RNA polymerase proteins.
RdDM has been implicated in a number of regulatory processes in plants. The DNA methylation added by RdDM is generally associated with transcriptional repression of the genetic sequences targeted by the pathway. Since DNA methylation patterns in plants are heritable, these changes can often be stably transmitted to progeny. As a result, one prominent role of RdDM is the stable, transgenerational suppression of transposable element (TE) activity. RdDM has also been linked to pathogen defense, abiotic stress responses, and the regulation of several key developmental transitions. Although the RdDM pathway has a number of important functions, RdDM-defective mutants in Arabidopsis thaliana are viable and can reproduce, which has enabled detailed genetic studies of the pathway. However, RdDM mutants can have a range of defects in different plant species, including lethality, altered reproductive phenotypes, TE upregulation and genome instability, and increased pathogen sensitivity. Overall, RdDM is an important pathway in plants that regulates a number of processes by establishing and reinforcing specific DNA methylation patterns, which can lead to transgenerational epigenetic effects on gene expression and phenotype.
Biological functions
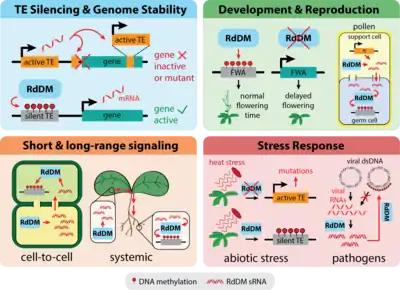
RdDM is involved in a number of biological processes in the plant, including stress responses, cell-to-cell communication, and the maintenance of genome stability through TE silencing.
Transposable element silencing and genome stability
TEs are pieces of DNA that, when expressed, can move around the genome through a copy-and-paste or cut-and-paste mechanism. New TE insertions can disrupt protein coding or gene regulatory sequences, which can harm or kill the host cell or organism.[1] As a result, most organisms have mechanisms for preventing TE expression. This is particularly key in plant genomes, which are often TE-rich. Some plant species, including important crops like maize and wheat, have genomes consisting of upwards of 80% TEs.[1][2] RdDM plays a key role in silencing these mobile DNA elements in plants by adding DNA methylation over new TE insertions and constantly reinforcing DNA methylation over existing TEs, inhibiting transposition and maintaining long-term genome stability.[3] Although the RdDM mechanism itself is unique to plants, using DNA methylation to silence TEs is a common strategy among eukaryotes.[4]
RdDM primarily targets small TEs and TE fragments near genes, which are usually in open, accessible euchromatic regions of the genome that are permissive for gene expression.[3][5] In these regions, the ‘active’ chromatin state has a tendency to spread from expressed genes to nearby repressed regions, like TEs, which can cause these TEs to become activated and transpose.[3] Continuous activity by RdDM opposes the spread of active chromatin, maintaining a silent, repressive heterochromatic state over TEs in these otherwise euchromatic regions. In turn, RdDM activity recruits other pathways that help establish and propagate the silent, heterochromatic state (see 'Interactions between RdDM and other chromatin modifying pathways'). Because of the self-reinforcing nature of these silencing pathways, excessive RdDM activity can also cause the silent, heterochromatic chromatin state over TEs to spread to nearby genes and repress them, with potentially harmful consequences for the organism.[3][5] Therefore, RdDM activity must be finely tuned to maintain a balance between repressing TEs and allowing expression of nearby genes.[3]
In addition to maintaining stable silencing of TEs, RdDM can also initiate transcriptional silencing of foreign DNA, including novel TE insertions, virus-derived sequences, and transgenes (also see 'Biotic stresses' and 'Transgene silencing' below).[6][7][8][9][10] When TEs integrate near genes, RdDM-mediated silencing of the TEs often affects gene expression.[3][1] However, this is not always deleterious, and can sometimes be overcome by other processes,[11] or alter gene expression in ways beneficial to the plant. Over evolutionary time, beneficial TEs can become an important part of the mechanism by which a gene is regulated.[3][1] In one example, the gene ROS1 lies adjacent to a small helitron TE that is normally methylated by RdDM.[12][13] While DNA methylation is normally associated with transcriptional repression, this is not the case at the ROS1 locus. Instead, methylation of the helitron TE promotes ROS1 expression, so ROS1 expression is lost in mutants of the RdDM pathway that cannot methylate the TE.[12][13] Interestingly, ROS1 encodes a DNA glycosylase that functions to remove DNA methylation from the genome.[14] The link between ROS1 expression and RdDM activity at this TE ensures that DNA methylation and demethylation activities remain in balance, helping to maintain DNA methylation homeostasis genome-wide.[12][13] Thus, RdDM-mediated regulation of TEs can lead to beneficial regulatory outcomes.
Some TEs have evolved mechanisms to suppress or escape RdDM-based silencing in order to facilitate their own proliferation, leading to an evolutionary arms race between TEs and their host genomes. In one example, a TE-derived sequence was found to produce sRNAs that trigger post-transcriptional repression of a component of the RdDM pathway, inhibiting RdDM.[15] This sequence may have helped the original TE escape RdDM-based silencing and insert itself into the host genome.
Studying how RdDM targets and represses different types of TEs has led to many major insights into how the RdDM mechanism works. The retrotransposon EVADÉ (EVD) was one of the first TEs specifically shown to be repressed by RdDM-derived sRNAs.[16] Later work used EVD to trace the mechanism by which a novel TE insertion became silenced, revealing an important mechanistic link between post-transcriptional gene silencing and RdDM.[9] Studies of other retrotransposons, including ONSEN, which is regulated by both RdDM and heat stress,[17][18] and Athila family TEs,[10] among many others, have also provided valuable insights into RdDM-mediated TE silencing.
Development and reproduction
A number of epigenetic changes required for normal development and reproduction in flowering plants involve RdDM. In a well-studied example, RdDM is required for repression of the FWA gene, which allows for proper timing of flowering in Arabidopsis.[19] The FWA promoter contains tandem repeats that are usually methylated by RdDM, leading to transcriptional repression.[20] Loss of this methylation re-activates FWA expression, causing a late-flowering phenotype.[19][20] The loss of DNA methylation and associated late-flowering phenotype can be stably transmitted to progeny. Since the demethylated fwa allele leads to a stable, heritable change in the expression of FWA without any change to the DNA sequence, it is a classic example of an epiallele.
Mutations in the RdDM pathway can strongly affect gamete formation and seed viability, particularly in plant species with high TE content like maize and Brassica rapa, highlighting the importance of this pathway in plant reproduction.[21][22][23] During gamete formation, it has been hypothesized, and in some cases shown, that RdDM helps reinforce TE silencing in the germ cells.[24][25] In both pollen and ovules, a support cell undergoes epigenetic reprogramming, losing DNA methylation and other epigenetic marks at a number of loci, including TEs.[26][24] This causes TE re-activation and encourages the production of RdDM-derived sRNAs against these TEs in the support cells. The sRNAs are then thought to move from the support cell to the germ cell in order to reinforce TE silencing in the next generation. This phenomenon has been observed in pollen, but has yet to be shown definitively in the ovule.[27][28] This role for sRNAs in plants resembles the role of piRNAs in germline development in Drosophila and some other animals.[29][30] A similar phenomenon may also occur in roots to preserve TE silencing in important stem cell populations.[31]
The RdDM pathway is also involved in regulating imprinted expression at some genes.[32] This unusual parent-of-origin-specific expression pattern occurs at several loci in the endosperm during seed development in flowering plants. A few factors involved in the RdDM pathway are themselves imprinted (favoring expression from the paternal allele) in diverse species, including A. thaliana, A. lyrata, C. rubella, and maize.[33][34][35][36] RdDM also plays a role in mediating the gene dosage effects seen in seeds derived from interploid crosses,[37][38] though the mechanism for this remains largely unknown.
There is also evidence that RdDM plays a role in several other aspects of plant development, including seed dormancy,[39] fruit ripening,[40] and other pathways involved in flowering.[41] However, most of these data are correlative, and further study is necessary to understand the role of RdDM in these processes.
Stress response
Abiotic stresses
RdDM helps plants respond to a number of abiotic stresses, such as heat stress, drought, phosphate starvation, salt stress, and others.[42] Many TEs become upregulated under abiotic stress conditions,[43][44] and thus one function of RdDM in stress response is to help counter this activation. In one example, the retrotransposon ONSEN is upregulated by heat stress, but normally remains suppressed by RdDM-associated sRNAs and can only transpose efficiently in heat-stressed plants that are also deficient in RdDM.[17][18] More generally, in plants exposed to heat stress, several components of the RdDM pathway become upregulated, and mutations in some components of the RdDM machinery reduce heat tolerance, suggesting RdDM plays an important role during heat stress.[45][46] In addition to regulating TEs under stress conditions, RdDM can also regulate genes in order to trigger appropriate stress responses. Under low humidity, leaves produce fewer stomata due to RdDM-mediated downregulation of two genes involved in stomatal development.[47] Similarly, RdDM becomes downregulated in response to salt stress, and this has been shown to trigger the expression of a transcription factor important in salt stress resistance.[48]
Biotic stresses
RdDM was initially discovered as a response to infection by viroids,[49] and along with RNAi plays an important role in defending the plant against viroids and viruses. The RdDM and RNAi machinery recognize viral RNAs and process them into sRNAs, which can then be used both pathways to degrade viral RNA (RNAi) and silence viral DNA (RdDM).[50][51][52] However, little is known about how the RdDM and RNAi machinery distinguish between viral RNAs and RNAs produced by the host plant. Mutants defective in RdDM and other methylation-deficient mutants are often hypersensitive to viral infection.[53][54] Virus-host interactions are another example of an evolutionary arms race, and many plant viruses encode suppressors of both RdDM and RNAi in an attempt to evade the host plant's defenses.[55][53][56][57]
RdDM is also involved in protecting the plant from other biotic stresses,[50] including bacterial infections,[58] fungal infections,[59] and predation.[60] Loss of RdDM can have opposing effects on resistance for different pathogens. For example, some RdDM mutants have increased susceptibility to the bacterium Agrobacterium tumefaciens,[61] but those same mutants have decreased susceptibility to the bacterium Pseudomonas syringae,[58] highlighting the complexity of the different pathogen defense pathways and their interactions with RdDM.[62]
Transgene silencing
In addition to naturally-occurring foreign nucleic acid stressors like TEs and viruses, artificially introduced DNA sequences, like transgenes, are also targeted for repression by RdDM.[63][6] Transgenes are widely used in genetics research to study gene function and regulation, and in plant breeding to introduce novel and desirable properties into a plant. Transgene silencing by RdDM and other mechanisms has therefore proved problematic for plant researchers. Efforts to understand how transgenes become silenced have ultimately helped reveal much of what we now know about the RdDM pathway (see 'History and discovery of RdDM'). In one early example, researchers sequentially transformed plants with two different transgenes that shared some of their DNA sequence.[64] They found that transforming the second transgene into the plants led to the first transgene gaining DNA methylation and becoming inactivated.[64] This provided an early clue that there existed a trans-acting, sequence-based mechanism for transcriptional silencing of foreign DNA, later shown to be RdDM.
Stress and RdDM-mediated epigenetic ‘memory’
Due to the heritability of DNA methylation patterns in plants, and the self-reinforcing nature of RdDM and other DNA methylation pathways, any DNA methylation changes caused by environmental stressors have the potential to be maintained and transmitted to future generations. This can allow stress-induced DNA methylation changes to act as a ‘memory’ of the stressor and help prime the plant or its progeny to respond more efficiently to the stress if re-exposed.[50][65] For example, RdDM-derived sRNAs against TEs or viruses that have already integrated into the genome and been silenced serve as a 'memory' of those prior infections, protecting against future invasions by similar sequences. There is also evidence that DNA methylation changes due to other stressors, such as salt or heat stress, can persist in the progeny of stressed plants even in the absence of the original stressor.[66] In this study, the persistence of the stress-induced DNA methylation changes required several RdDM-related proteins, suggesting that RdDM was involved in maintaining the stress-altered DNA methylation patterns. In another example, resistance to insect attack was transmitted to progeny via DNA methylation changes, and this inheritance was also dependent on functional sRNA biogenesis pathways.[60][50] Thus, RdDM can potentially alter the plant epigenome in response to stress, and helps maintain these changes to modulate future stress responses in the affected plant and its descendants.
Short and long-range signaling
The sRNA molecules produced by RdDM and other pathways are able to move between cells via plasmodesmata, and can also move systemically through the plant via the vasculature.[67][68][69] They therefore have the potential to act as signaling molecules. This has been demonstrated in plants engineered to express green fluorescent protein (GFP).[70] The GFP protein produced by these plants caused them to glow green under certain light conditions. When tissue from a second plant expressing a sRNA construct complementary to GFP was grafted onto the GFP-expressing plant, the GFP fluorescence was lost: after grafting, the sRNAs being produced in the second plant's tissues were moving into the tissues of the first, GFP-expressing plant, and triggering silencing of GFP.[70] The same study showed that a subset of these mobile sRNAs were triggering the addition of DNA methylation to the GFP locus via RdDM. Therefore, sRNAs involved in RdDM can act as signaling molecules and trigger the addition of DNA methylation at complementary loci in cells far away from where the sRNAs were originally generated. Since then, studies have shown that sRNAs can move and direct RdDM both from shoot to root and root to shoot, though the silencing effect is more robust when sRNAs move from shoot to root.[69][70][71][72]
Movement of sRNAs that drive RdDM activity plays an important role in plant development, including during reproduction[23][24][27] and root development.[31] In both cases, sRNA movement seems to function primarily as a way to reinforce DNA methylation and silencing of TEs in developmentally important cell types, like germ cells and stem cells. Silencing TEs and maintaining genome integrity in these cells is particularly important because they give rise to many other cells, all of which will inherit any defects or mutations in the original stem cell or germ cell. sRNA movement is also involved in plant-pathogen interactions: sRNAs can move from infected cells to distal uninfected tissues in order to prime a defense response, though to date this has only been shown for RNAi, not RdDM.[73]
Pathways and mechanisms
This section focuses on the pathways and mechanisms by which RdDM leads to sequence-specific DNA methylation. The pathways presented here were characterized primarily in the model plant Arabidopsis thaliana, but are likely similar in other angiosperms. Conservation of RdDM in other plant species is discussed in more detail in 'Evolutionary conservation' below.
DNA methylation context
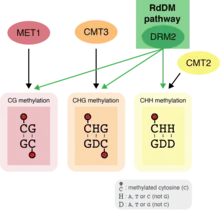
RdDM is the only mechanism in plants that can add DNA methylation to cytosines regardless of sequence context.[55] DNA methylation in plants is typically divided into three categories based on the sequence context of the methylated cytosine: CG, CHG, and CHH, where H is any nucleotide except G. These reflect the different sequence contexts targeted by several DNA methylation pathways in plants. These context-specific pathways are primarily involved in maintaining existing DNA methylation patterns. The highly conserved methyltransferase MET1 (homolog of mammalian DNMT1) maintains DNA methylation in the CG context, while two conserved plant-specific methyltransferases, Chromomethylase 3 (CMT3) and CMT2, help maintain CHG and CHH methylation, respectively.[74][75][76][77] Unlike these pathways, RdDM leads to the addition of DNA methylation at all cytosines regardless of their sequence context. Like MET1, CMT2 and CMT3, RdDM is primarily involved in maintaining existing DNA methylation patterns.[55] However, RdDM is also the only pathway capable of adding DNA methylation de novo to previously unmethylated regions in plants.
Mechanism
The RdDM pathway can be split up into two main processes: the production of sRNAs, and the recruitment of DNA methylation machinery by those sRNAs to specific target loci in the DNA.[78][55][79] These two activities together constitute RdDM, and ultimately lead to DNA methylation being added to cytosines at specific target loci.
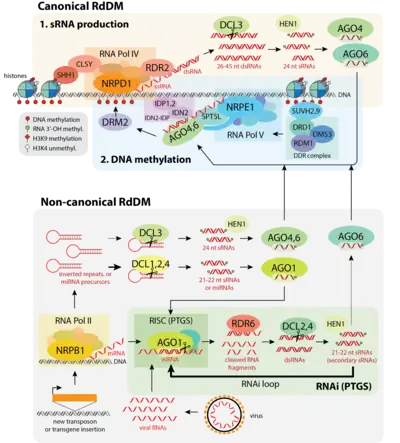
Canonical RdDM
The canonical RdDM pathway is, as its name suggests, the most well-characterized RdDM pathway to date. Canonical RdDM is preferentially recruited to regions that are already DNA-methylated and heterochromatic, and acts to reinforce existing DNA methylation patterns at these loci, forming a positive feedback loop.[55][79] Canonical RdDM makes up the majority of RdDM activity in a cell.[79]
sRNA production
The first part of the RdDM pathway revolves around the biogenesis of sRNAs. A plant-specific RNA polymerase complex, RNA Polymerase IV (Pol IV), is first recruited to silent heterochromatin via its interaction with CLASSY (CLSY) proteins and SAWADEE homeodomain homolog 1 (SHH1) (also see 'Interactions between RdDM and other chromatin modifying pathways' below).[80][79][81] Pol IV transcribes these regions to produce short single-stranded RNAs (ssRNAs) roughly 30 to 45 nucleotides in length, each of which is the precursor for a single sRNA.[82][83][84] These ssRNAs are converted into double-stranded RNAs (dsRNAs) co-transcriptionally by RNA-directed RNA polymerase 2 (RDR2), which physically associates with Pol IV.[83] The dsRNAs are then cleaved by the endoribonuclease Dicer-like 3 (DCL3) into 24 nucleotide (nt) sRNAs. Pol IV, RDR2, and DCL3 alone are sufficient for the production of 24 nt sRNAs in vitro,[84] suggesting that while other factors involved in this part of the pathway may help increase efficiency or specificity, they are not required for Pol IV-mediated sRNA production.
While nearly all 24 nt sRNAs involved in RdDM are produced through the Pol IV-RDR2-DCL3 pathway, a small proportion are produced through other pathways. For example, some RNA Polymerase II (Pol II) transcripts that contain an inverted repeat sequence form double-stranded hairpin structures that can be directly cleaved by DCL3 to form 24 nt sRNAs.[85][79]
DNA methylation of target loci
In the second part of the pathway, the RdDM DNA methylation machinery is guided to DNA sequences complementary to the sRNAs generated in the first part of the pathway. One strand from each 24 nt double-stranded sRNA is loaded into Argonaute (AGO) proteins AGO4, AGO6, or AGO9.[55] AGO3 may also be able to function in this pathway.[86] Argonautes are a large, highly conserved family of proteins that can bind sRNAs, forming a protein-sRNA duplex that enables them to recognize and bind other RNA sequences complementary to their sRNA partner.[87] Once formed, the AGO-sRNA duplex finds and binds complementary sequences along an RNA ‘scaffold’ produced by the plant-specific RNA polymerase V (Pol V), with the help of interactions with Suppressor of Ty insertion 5-like (SPT5L), the Involved in de novo 2 - IDN2 Paralog (IDN2-IDP) complex, and the Pol V subunit NRPE1.[88] This leads to the recruitment of the DNA methyltransferase enzyme Domains Rearranged Methyltransferase 2 (DRM2), which methylates nearby DNA.[89][55][79] The mechanism by which the AGO-sRNA duplex recruits DRM2 is not yet well understood.[90]
Non-canonical RdDM
Recent work has revealed a number of variations of the RdDM pathway, collectively referred to as non-canonical RdDM.[79] Unlike canonical RdDM, the non-canonical pathways are generally involved in establishing initial DNA methylation at new target loci, like novel TE insertions, rather than maintaining existing heterochromatin. Actively expressing elements like new TE insertions are normally strongly targeted by post-transcriptional gene silencing (PTGS/RNAi) pathways. Non-canonical RdDM occurs primarily as a byproduct of these PTGS pathways, leading to the initial establishment of a silent, heterochromatic state over the new TE or other target locus. Once that initial silent state is established, Pol IV can be recruited to the locus by CLSY and SHH1, and the canonical RdDM pathway takes over the long-term maintenance of silencing.[79] Therefore, the non-canonical RdDM pathways often act as a temporary bridge between initial post-transcriptional silencing of novel elements by RNAi, and long-term transgenerational transcriptional silencing via canonical RdDM.[10][9][79] Consistent with this role in initiation of novel silencing, non-canonical RdDM targets relatively few loci in comparison to canonical RdDM.[79]
The primary difference between the canonical and non-canonical RdDM pathways lies in the origin and biogenesis of the sRNAs involved. The canonical RdDM pathway involves 24 nt sRNAs, which are specific to that pathway and come predominantly from a single source (the Pol IV-RDR2 complex). In contrast, the non-canonical RdDM pathways involve 21-22 nt sRNAs from a variety of sources, allowing de novo DNA methylation to be initiated at many different types of loci. These 21-22 nt sRNAs are not specific to non-canonical RdDM, and also function in other PTGS pathways. In fact, only a small fraction of 21-22nt sRNAs are involved in RdDM, with the majority instead driving a positive feedback loop amplifying the PTGS response.[91] The functional outcome of a specific 21-22 nt sRNA depends on the AGO protein it ultimately associates with: sRNAs that associate with AGO4, AGO6 or AGO9 result in RdDM and DNA methylation, while sRNAs that associate with other AGOs, like AGO1, primarily result in PTGS.[55][79]
By using 21-22 nt sRNAs derived from a variety of sources, non-canonical RdDM can flexibly induce de novo DNA methylation and silencing at many different types of loci. One of the primary sources of 21-22 nt sRNAs is Pol II transcripts. Some of these transcripts, particularly those produced from TEs, viruses, or certain non-protein-coding transcripts, are targeted by PTGS pathways like miRNAs or RNAi, leading to cleavage of the transcript. The resulting fragments can be converted into dsRNA by RDR6 and then processed into 21-22 nt sRNAs by DCL2 or DCL4.[8] Most of these 21-22 nt sRNAs are loaded into AGO1 and feed back into PTGS, amplifying PTGS efficiency.[79] However, some will instead associate with AGO6, leading to RdDM.[10] dsRNAs resulting from RDR6 activity can also sometimes processed by DCL3 instead of DCL2/4 and trigger RdDM.[9] Additionally, some Pol II transcripts contain inverted repeat sequences, which can form double-stranded hairpin-like structures. These can be cleaved by DCL proteins independent of RDRs to produce either 21-22 nt or 24 nt sRNAs that can participate in RdDM.[79] Similarly, miRNA precursors, which also form hairpin structures and are normally cleaved by DCL1 to produce miRNAs, can instead be cleaved by other DCLs to form sRNAs for RdDM.[79] While most non-canonical RdDM occurs via AGO6 or AGO4, there is also a version of the pathway where sRNAs instead associate with AGO2, which together with the NERD complex (Needed for RDR2-independent DNA methylation) recruits DRM2 to target loci and triggers DNA methylation.[92] Since the non-canonical pathways are not yet as well characterized as the canonical RdDM pathway,[79] there likely remain additional sources of sRNAs used for RdDM that have not yet been uncovered.
Factors involved
A number of factors involved in RdDM are listed below, along with additional details about their function and corresponding references. Several factors primarily involved in PTGS that sometimes participate in RdDM are also listed.
| Factor(s) | Factor type | Pathway | Role in RdDM | Known direct interactors | Description | References |
|---|---|---|---|---|---|---|
| NRPD1 and the Pol IV complex | RNA polymerase | Canonical RdDM | sRNA production | CLSY proteins, RDR2 | Pol IV is a plant-specific RNA polymerase complex and NRPD1, its largest subunit, is specific to the complex. Through its interaction with the CLSY proteins and SHH1, Pol IV is recruited to heterochromatic regions (specifically to H3K9me2- and H3K4me0-containing chromatin), and transcribes single-stranded RNAs precursors of the sRNAs used in the canonical RdDM pathway. | [93][80][94][81] |
| NRPE1 and the Pol V complex | RNA polymerase | All RdDM | DNA methylation of target loci | Pol V is a plant-specific RNA polymerase complex and NRPE1, its largest subunit, is specific to the complex. Pol V transcribes non-coding RNAs that serve as scaffolds for several other RdDM components, most importantly the AGO-sRNA duplex, but also SPT5L, and the IDN2-IDP complex. Both NRPE1 and SPT5L contain an AGO hook motif that helps recruit AGO4 to Pol V transcripts. Mutating the AGO hook motifs on both proteins results in reduced DNA methylation at RdDM target loci, resembling nrpe1 null mutant phenotypes. Binding of the AGO-sRNA duplex to complementary sites along the Pol V transcript leads to recruitment of DRM2 and addition of DNA methylation to target loci. | [93][80][94][95][81] | |
| RDR2 | RNA-dependent RNA polymerase | Canonical RdDM | sRNA production | Pol IV | Exists in a complex with Pol IV and converts the nascent Pol IV transcript to double-stranded RNA, which can then be processed by DCL3 to generate sRNAs for canonical RdDM. | [83][80] |
| RDR6 | RNA-dependent RNA polymerase | PTGS, non-canonical RdDM | sRNA production | Converts single-stranded RNAs to double-stranded RNAs for processing into 21-22 nt sRNAs by DCL2 and DCL4. Most of these sRNAs lead to PTGS, but some are loaded into AGO6 and participate in non-canonical RdDM. | [9][80] | |
| DCL1 | Endoribonuclease | PTGS, non-canonical RdDM | miRNA production, sRNA production | An endoribonuclease that cleaves double-stranded RNA, primarily involved in the production of microRNAs that lead to PTGS via AGO1. Can also catalyze the production of 21 nt sRNAs from mRNAs containing inverted repeats, which can be used in either PTGS or non-canonical RdDM depending on the AGO protein they associate with. The four DCL proteins in A. thaliana (DCL1,2,3,4) compete for access to dsRNA substrates. | [96][97][81][98] | |
| DCL2 | Endoribonuclease | PTGS, Non-canonical RdDM | sRNA production | An endoribonuclease that cleaves double-stranded RNA, resulting in 22 nt sRNAs that can be used in both PTGS and non-canonical RdDM. The four DCL proteins in A. thaliana (DCL1,2,3,4) compete for access to dsRNA substrates, and DCL2,4 can substitute for loss of DCL3 for most RdDM targets. | [96][99][97][80] | |
| DCL3 | Endoribonuclease | Canonical RdDM | sRNA production | An endoribonuclease that cleaves double-stranded RNA, resulting in 24 nt sRNAs used in canonical RdDM. Preferentially targets the short dsRNAs produced by Pol IV-RDR2, but can also slice other dsRNA substrates, including mRNAs containing inverted repeats or miRNA precursors. The four DCL proteins in A. thaliana (DCL1,2,3,4) compete for access to dsRNA substrates, and DCL2,4 can substitute for loss of DCL3 for most RdDM targets. When PTGS pathways via DCL2,4 become saturated, DCL3 can step in and process the DCL2,4 dsRNA substrates, triggering a switch from PTGS to RdDM-mediated TGS. | [96][9][99][97][80] | |
| DCL4 | Endoribonuclease | PTGS, Non-canonical RdDM | sRNA production | An endoribonuclease that cleaves double-stranded RNA, resulting in 21 nt sRNAs that can be used for both PTGS and non-canonical RdDM. The four DCL proteins in A. thaliana (DCL1,2,3,4) compete for access to dsRNA substrates, and DCL2,4 can substitute for loss of DCL3 for most RdDM targets. | [96][99][97] | |
| AGO4 | Argonaute protein | Canonical RdDM | DNA methylation of target loci | NRPE1, SPT5L | The main Argonaute protein involved in canonical RdDM. AGO4 is partially redundant with AGO6, which can also function in this pathway, as well as with AGO9 in reproductive tissues. It binds the 24 nt sRNAs produced by the pathway to form an AGO4-sRNA duplex, which can recognize sequences complementary to the sRNA. Assisted by interactions with SPT5L, NRPE1, and the IDN2-IDP complex, the AGO4-sRNA duplex binds a single-stranded, noncoding RNA produced by Pol V, and helps recruit DRM2 to the DNA. | [93][80][100] |
| AGO6 | Argonaute protein | All RdDM | DNA methylation of target loci | An argonaute protein that can function in either canonical or non-canonical RdDM pathways. Partially redundant with AGO4 (the main canonical RdDM AGO). Can associate with either 24 nt or 21-22 nt sRNAs to trigger RdDM at complementary loci. By interacting with both 21-22 nt and 24 nt sRNAs, AGO6 helps in the transition from PTGS (normally mediated by 21-22 nt sRNAs) to stable silencing by RdDM (normally mediated by 24 nt sRNAs). Expressed particularly in the root and shoot meristems, which are the two main stem cell populations in plants. This may indicate that plants increase surveillance for novel TEs in order to ensure genome integrity in the key cells that will give rise to most of the other cells in the plant. | [93][101][10][80][100] | |
| AGO9 | Argonaute protein | Canonical RdDM | DNA methylation of target loci | A highly specialized AGO expressed primarily in the germline, where it is required for proper female gamete formation. Interacts with 24 nt sRNAs to silence TEs in the germline, similar to the role of PIWI Argonaute proteins in animals. | [102][25][100] | |
| AGO1 | Argonaute protein | PTGS, non-canonical RdDM | sRNA production | Binds microRNAs or 21-22 nt sRNAs, which it uses to recognize complementary sequences on other RNAs. When an AGO1-sRNA duplex (often called the RISC) finds a complementary single-stranded mRNA, the RNA is cleaved by AGO1, destroying the mRNA and causing PTGS. The resulting RNA fragments can then be converted to dsRNAs by RDR6 and processed by DCL2,4 to form secondary 21-22 nt sRNAs. These are predominantly loaded back into AGO1, forming a self-reinforcing ‘RNAi loop’. However, some of the 21-22 nt sRNAs are loaded into AGO6 instead, leading to RdDM. | [91][97][80][100] | |
| DRM2 | DNA methyltransferase | All RdDM | DNA methylation of target loci | The main DNA methyltransferase involved in RdDM. Catalyzes the addition of a methyl group to cytosines in DNA. Recruited by the AGO4-sRNA duplex after it binds to a complementary sequence in a Pol V transcript, but the mechanism by which this happens is not well understood. | [103][80] | |
| SHH1/DTF1 | DNA and chromatin binding protein | Canonical | sRNA production | CLSY1 | Required for Pol IV-derived sRNA production at a subset of RdDM loci. Via its SAWADEE domain, SHH1 binds histone H3 with specific modifications associated with heterochromatin and DNA methylation: methylation of the 9th lysine (H3K9me2) and unmethylated K4 (H3K4me0). By interacting with SHH1 via the CLSY proteins, Pol IV is recruited to heterochromatic/silent chromatin. To date, SHH1 has only been shown to directly interact with CLSY1. The ability of SHH1 to associate with Pol IV/NRPD1 is mostly abolished in clsy1,2 double mutants, so recruitment of Pol IV by SHH1 likely requires CLSY proteins. | [104][105][106][107] |
| CLSY1, CLSY2 | putative chromatin remodelers | Canonical | sRNA production | Pol IV, SHH1 | Required for SHH1 interaction with and recruitment of Pol IV to a subset of target loci. Mutually exclusive with loci regulated by CLSY3 and CLSY4. Together, the four CLSY proteins regulate nearly all Pol IV-derived sRNAs, and loss of all four results in a near total loss of 24-nucleotide sRNA production. Requires H3K9me2, likely through interaction with SHH1. sRNAs regulated by CLSY1,2 are enriched in the chromosome arms, while those regulated by CLSY3,4 are enriched in the pericentromere. | [107][108] |
| CLSY3, CLSY4 | putative chromatin remodelers | Canonical | sRNA production, Pol IV targeting | Pol IV | Involved in recruitment of Pol IV to a subset of target loci. Mutually exclusive with loci regulated by CLSY1 and CLSY2. Together, the four CLSY proteins regulate nearly all Pol IV-sRNAs, and loss of all four results in a near total loss of 24-nucleotide sRNA production. sRNAs regulated by CLSY3,4 are enriched in the pericentromere, while sRNAs regulated by CLSY1,2 are enriched in the chromosome arms. | [107][108] |
| HEN1 | RNA methylase | Both | sRNA production | none | Stabilizes sRNAs by adding methylation to the 3'-OH groups. | [109] |
| SUVH2, SUVH9 | methyl-DNA binding proteins | Both | DNA methylation of target loci | DDR complex, MORC1, MORC6 | A pair of closely related methyl-DNA binding proteins that interact with the DDR complex and are required for proper localization of the DDR complex and Pol V. By recruiting Pol V to regions with DNA methylation, which tend to be silent, heterochromatic regions, SU(VAR)3-9 homolog (SUVH) 2 and 9 help form a positive feedback loop that reinforces RdDM-mediated silencing. May also associate with MORCs. | [110] |
| DDR complex (RDM1, DMS3, DRD1) | putative chromatin remodeling complex | Both | DNA methylation of target loci | SUVH2, SUVH9 | The DDR complex, composed of DRD1, DMS3, and RDM1, is thought to facilitate access of Pol V to its target sites, possibly by unwinding DNA downstream of Pol V. Interacts with SUVH2,9, which bind methylated DNA, and this interaction may help recruit Pol V to regions of existing heterochromatin. RDM1 also binds single-stranded DNA, which may help unwind the DNA to facilitate recruitment of DRM2. | [88][111][112][113][110] |
| SPT5L/RDM3/KTF1 | transcription factor | Both | DNA methylation of target loci | AGO4, Pol V transcripts | Interacts with AGO4 and helps recruit it to the RNA scaffold produced by Pol V. Like the Pol V subunit NRPE1, SPT5L contains an AGO hook motif in its C-terminal domain. The motifs on both NRPE1 and SPT5L redundantly help recruit AGO4 to loci being transcribed by Pol V. Mutating the AGO hook motifs on both proteins results in reduced DNA methylation at RdDM target loci, resembling nrpe1 null mutant phenotypes. Also required for co-transcriptional slicing of Pol V transcripts. | [114][95][115] |
| SWI/SNF complex | chromatin remodeling complex | Both | DNA methylation of target loci | IDN2 | The Switch/Sucrose non-fermentable (SWI/SNF) complex is a chromatin remodeling complex that is recruited to Pol V scaffolds by the IDN2-IDP complex, where it affects nucleosome positioning. SWI/SNF may promote RdDM by making the chromatin more accessible, which may facilitate access of DRM2 to DNA. | [116] |
| IDN2-IDP complex | dsRNA-binding protein | Both | DNA methylation of target loci | SWI/SNF complex | A complex composed of IDN2 and IDP1 (also called IDNL1) or IDP2 (IDNL2). IDN2, and possibly IDP1, can bind the dsRNA duplex formed when AGO-associated sRNAs hybridize with the Pol V scaffold. This complex is thought to help stabilize base pairing between the AGO-sRNA and Pol V scaffold RNA. IDN2-IDP may also facilitate recruitment of the SWI/SNF complex to Pol V scaffolds. Additionally, IDP1 can bind unmethylated DNA, which may help recruit DRM2 to regions lacking DNA methylation. | [117][116][118] |
| NERD | GW repeat- and PHD finger-containing protein | Non-canonical RdDM | sRNA production, DNA methylation of target loci | AGO2 | Forms a non-canonical RdDM pathway that includes a number of genes involved in PTGS, including AGO2. Binds histone H3 and AGO2. Required for 21 nt sRNA accumulation at some non-canonical RdDM targets, including novel TE insertions. Leads to histone tail modifications associated with transcriptional repression; because these modifications can recruit other DNA methylation machinery, including canonical RdDM, it is unclear if the effect of NERD on DNA methylation is direct or indirect. | [92][79] |
| MORC1, MORC6 | GHKL ATPases | Both | DNA methylation of target loci (?) | SUVH2, SUVH9, IDN2, DMS3 | Microrchidia 1 (MORC1) and MORC6 form a heterodimer and may interact with the DDR complex to recruit Pol V. However, they are thought to mainly act downstream of DNA methylation to promote silencing. Their precise role in RdDM is still unclear. | [110][80][90] |
| DRM1 | DNA methyltransferase | All RdDM | DNA methylation of target loci | A homolog of DRM2 that is only expressed during sexual reproduction, specifically in the egg cell and potentially the early embryo. DRM2 is likely the main RdDM methyltransferase in all other tissues. | [119] | |
| HDA6 | Histone deacetylase | Canonical RdDM | sRNA production | May facilitate Pol IV recruitment by creating a permissive chromatin state for SHH1 binding by removing histone acetylation, promoting H3K9 methylation. In histone deacetylase 6 (hda6) mutant plants, HDA6 target loci lose Pol IV targeting and sRNA biogenesis, suggesting HDA6 is involved in Pol IV recruitment at a subset of RdDM target loci. Further, normal Pol IV targeting cannot be restored after re-introduction of functional HDA6, suggesting that HDA6 is also required to propagate the trans-generational 'memory' of where Pol IV should be targeted. HDA6 physically associates with MET1 and facilitates CG methylation maintenance by MET1, which may also be important for sRNA production at HDA6-dependent loci. | [120][80] | |
Interactions with other chromatin modifying pathways
Different chromatin states, like active euchromatin or silent heterochromatin, are defined by a combination of specific histone modification and DNA methylation patterns. Repressive chromatin modifications, like DNA methylation, help promote DNA compaction and reduce DNA accessibility, while other modifications help open chromatin and increase accessibility. Methylation of the 9th lysine of histone H3 (H3K9), primarily in the form of H3K9 trimethylation (H3K9me3) in animals and H3K9 dimethylation (H3K9me2) in plants, is a highly conserved repressive modification.[121][122] Lack of H3K4 methylation (H3K4me0) is also associated with repression, along with several other histone modifications and variants. The combination of DNA methylation, H3K9me2, and H3K4me0 is strongly associated with heterochromatin in plants.
Since DNA methylation and repressive histone modifications together define heterochromatin, most DNA methylation pathways in plants recognize and interact with repressive histone marks and vice versa, forming positive feedback loops that help maintain the repressive chromatin state.[123] The RdDM-associated protein SHH1 recognizes H3K4me0 and H3K9me2 at heterochromatic loci and recruits Pol IV to these loci to trigger additional DNA methylation at these regions.[106] Similarly, SUVH2 and SUVH9 help recruit Pol V to loci with DNA methylation.[110] Thus, both major parts of the canonical RdDM pathway are preferentially recruited to regions that are already in the silent, heterochromatic state marked by DNA methylation, H3K9me2, and H3K4me0. DNA methylation at these same heterochromatic loci is also recognized by the histone methyltransferases SUVH4/KYP, SUVH5, and SUVH6, which bind to non-CG methylation and add H3K9me2 to nearby histones,[123][124] closing the positive feedback loop. Similarly, CMT3 and CMT2, the two DNA methyltransferases involved in the maintenance of CHG and CHH methylation respectively,[75] both bind and add DNA methylation to H3K9me2-marked heterochromatin, forming their own feedback loop with SUVH4/5/6.[125][123] These interactions help strongly reinforce silencing at TEs and other heterochromatic regions.
A similar feedback loop occurs in animals. HP1 plays a vital role in maintaining heterochromatin by propagating H3K9 methylation through a positive feedback loop with the H3K9 methyltransferase SUV39H.[126] H3K9 methylation recruits HP1, which recruits SUV39H to deposit more H3K9 methylation.[126] Though HP1 is conserved in plants, its function in this feedback loop is not.[127] Instead, the positive feedback loops between H3K9me2 and the RdDM and CMT2/3 DNA methylation pathways fulfill a similar function in propagating H3K9me2. More recently, a plant-specific protein, Agenet Domain Containing Protein 1 (ADCP1), was also identified that may function analogously to HP1 in maintaining H3K9me2 levels in heterochromatin, facilitating heterochromatin formation.[128]
Ultimately, the constant reinforcement of silencing chromatin modifications at heterochromatic loci creates a repressive chromatin state wherein the DNA and histones (nucleosomes) become tightly packed together. This helps silence gene expression by physically inhibiting access to the DNA, preventing RNA Polymerase II, transcription factors and other proteins from initiating transcription.[129] However, this same compaction also prevents factors involved in heterochromatin maintenance from accessing the DNA, which could lead to the silent, compact state being lost. This is particularly true in the dense constitutive heterochromatin surrounding the centromere. In these regions, the chromatin remodeler DDM1 plays a crucial role in DNA methylation maintenance by displacing nucleosomes temporarily to allow methyltransferases and other factors access the DNA.[130][131][5] However, since most RdDM targets are small TEs in open, accessible and gene-rich regions (see “TE silencing and genome stability”), few RdDM sites require DDM1.[5][99] In fact, dense heterochromatin inhibits RdDM.[5] By contrast, CMT2 and CMT3 preferentially function in constitutive heterochromatin and depend strongly on DDM1 to maintain silencing over these regions.[131][5][3] Similarly, MET1, which maintains DNA methylation at CG sites after replication, requires DDM1 to access heterochromatin and maintain CG methylation in those regions.[132] Thus, DDM1 is a key regulator of DNA methylation in dense heterochromatin, but regulates sites mostly independently from RdDM.[5][99]
Interactions between RdDM and the other three maintenance DNA methylation pathways are limited and predominantly indirect. The DNA methyltransferase MET1 robustly maintains CG methylation genome-wide, including at RdDM target sites. In RdDM mutants, non-CG methylation at RdDM target sites is lost, but CG methylation is still maintained, suggesting that MET1 activity is independent of RdDM.[99] However, although met1 mutants lose CG methylation as expected, they also lose much of their non-CG methylation, including at RdDM target loci.[99] At these sites, silencing can still be initiated by RdDM in met1 mutants, but it is not maintained or transmitted to progeny, suggesting that MET1 is important for the maintenance, but not initiation, of silencing at a subset of RdDM target loci.[133][120] This effect is likely indirect: loss of MET1 leads to loss of H3K9me2 at some sites, which inhibits the recruitment of Pol IV and therefore prevents maintenance of DNA methylation via canonical RdDM, although the non-canonical pathways (which do not involve Pol IV) are not affected.[99][120] Loss of the histone deacetylase HDA6, which facilitates maintenance methylation by MET1 at some loci, has a similar effect, suggesting that multiple different factors involved in maintaining heterochromatin likely facilitate RdDM-mediated DNA methylation maintenance.[120]
Loss of RdDM leads to strong loss of non-CG methylation at TEs in gene-rich regions in the chromosome arms, but has little effect on DNA methylation levels in the constitutive heterochromatin around the centromere.[99][5][3] This suggests that CMT2 and CMT3, which function primarily to maintain CHG and CHH methylation in dense constitutive heterochromatin, do not depend on RdDM activity.[99][5][3] Similarly, in cmt2,cmt3 double mutants, many TEs in the chromosome arms remain methylated, presumably due to the persistent activity of RdDM, indicating that loss of CMT2/3 has little effect on RdDM activity.[5][3] This suggests that RdDM and CMT2/3 function mostly independently and at distinct loci: RdDM is the main pathway responsible for maintaining non-CG DNA methylation in euchromatic, gene rich regions, while CMT2 and CMT3 maintain non-CG DNA methylation in constitutive heterochromatin. In mutants defective in both RdDM and CMT2/CMT3, all non-CG methylation in the genome is eliminated,[74] demonstrating that together RdDM and CMT2/CMT3 account for all non-CG methylation in the genome.
Balance between DNA methylation and demethylation
Most DNA methylation mechanisms in plants are self-reinforcing (see above), including RdDM: Pol IV and Pol V are both recruited to heterochromatic regions that already have DNA methylation, encouraging additional DNA methylation via canonical RdDM.[55] Positive feedback loops like these can cause DNA methylation activity to spread out from the intended methylated target sites into genes or other regulatory elements, which can negatively affect gene expression. To prevent this spreading, DNA methylation pathways are opposed by passive and active DNA demethylation. DNA methylation can be lost passively with each cell division, because newly synthesized strands of DNA lack DNA methylation until it is re-added by one of the maintenance DNA methylation pathways.[134] DNA methylation can also be actively removed in plants by DNA glycosylases, which remove methylated cytosines via the base excision repair pathway. In Arabidopsis, there are four proteins responsible for removing DNA methylation: Repressor of silencing 1 (ROS1), Demeter (DME), Demeter-like 2 (DML2), and Demeter-like 3 (DML3).[135][136] These DNA glycosylases help prevent the spread of DNA methylation from RdDM targets to active genes.[137][14] Loss of active DNA demethylation in ros1;dml2;dml3 triple mutants leads to a widespread increase in DNA methylation levels, whereas ectopic expression of ROS1 leads to progressive loss of DNA methylation at many loci,[138] highlighting the importance of balancing DNA methylation and demethylation activity.
Interestingly, expression of the DNA demethylase ROS1 is directly tied to RdDM activity: DNA methylation over a TE targeted by RdDM in the ROS1 promoter is required for ROS1 expression,[12][13] though other factors are also involved in regulating ROS1.[139][140] Since ROS1 expression is tied to DNA methylation at a specific TE, ROS1 expression is also strongly reduced in plants with defective RdDM that lose the ability to methylate that TE and others.[12] This general mechanism helps maintain DNA methylation homeostasis by tuning DNA demethylation activity to DNA methylation activity, helping to ensure that DNA methylation patterns can be stably maintained over time.
Evolutionary conservation
Origins of RdDM pathway members
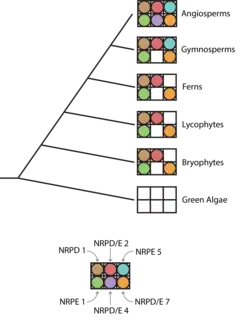

While all eukaryotes share three RNA polymerases (RNA Pol I, II and III), plants have two additional polymerases, Pol IV and Pol V. Both Pol IV and V share an evolutionary origin, deriving from Pol II.[141][94] In other eukaryotic kingdoms that lack these two specialized RNA polymerases, Pol II transcribes the precursors of small RNAs used in silencing pathways – in fact, Pol II transcripts are also sometimes processed into sRNAs in plants. It has been hypothesized that the origin of both Pol IV and Pol V is rooted in “escape from adaptive conflict”.[142] The idea is that potential tensions between the “traditional” function of Pol II and the small RNA biogenesis function could be relieved by duplication of Pol II and subfunctionalization of the resulting multiple RNA polymerases.
Analyses of evolutionary lineage for Pol IV and Pol V are complicated to some extent by the fact that each enzyme is actually composed of at least 12 subunits.[141] In Arabidopsis thaliana, some subunits are shared between Pol IV and Pol V, some are unique to each polymerase, and some are shared between Pol II, IV, and V.[143] Orthologs of certain Pol IV and V subunits have been found in all lineages of land plants, including ferns, liverworts, and mosses.[144][142] These findings argue for a shared origin of Pol IV and V dating back to early land / vascular plants.
Much of the work done to elucidate the genes and proteins involved in the RdDM pathway has been performed in Arabidopsis thaliana, a model angiosperm. However, studies of Pol IV and V conducted in maize show some key differences with Arabidopsis. Maize Pol IV and V differ from each other in terms of only one subunit (the largest one). In Arabidopsis, Pol IV and V differ from each other in terms of three subunits.[145] However, maize utilizes a set of interchangeable catalytic subunits – two in the case of Pol IV and three in the case of Pol V – that provide additional specialization of polymerase functionality.[145] While differences exist, overall there is a broad overlap in RdDM functions and components between the different angiosperm species studied to date.
Outside of Pol IV and Pol V, a large proportion of key RdDM component proteins (for example, DCL3 and AGO4) have orthologs found within each class of land plants, which provides support for the hypothesis that some form of the RdDM pathway evolved early within the plant lineage.[142] However, RdDM pathway functionality does appear to change to an appreciable extent between different plant species and lineages. For example, while gymnosperms have functional Pol IV and produce 24 nt small RNAs, the biogenesis of sRNAs within gymnosperms is much more heavily skewed towards 21 nt than 24 nt sRNAs.[146] This suggests that canonical RdDM may be rarer or less pronounced in gymnosperms than in angiosperms. Similarly, while orthologs of DRM2 are found in various angiosperms, there are no known DRM2 orthologs in other plant lineages.[147] One possibility is that angiosperms have the “most complete” version of the RdDM pathway, with all other plant lineages possessing robust and functional subsets of the pathway. However, since nearly all of the work on RdDM has been done in angiosperms, it is also possible that alternative versions of RdDM in other lineages have simply not yet been uncovered, particularly if these alternative versions include different proteins or proteins without clear homologs in angiosperms.
Relationships with sRNA silencing pathways in other kingdoms
All eukaryotic kingdoms host some form of small RNAs. One such class of sRNAs is the Piwi-interacting RNAs (piRNAs). Much like in RdDM, piRNAs primarily function to target and silence transposons, particularly in the germline.[29][30] However, piRNAs are only found in animals, are longer than the small RNAs functioning in RdDM (24-32 nucleotides), and mediate their functions through interactions with a different subclass of AGO proteins, the PIWI subfamily, which are absent from plants.[29][30] MicroRNAs (miRNAs) are another class of small RNA with silencing properties.[148] While miRNAs are in a similar size range as RdDM sRNAs (~21 nt), miRNAs associate with a distinct set of Argonaute proteins that silence target RNAs by initiating their degradation or blocking their downstream translation into proteins, rather than recruiting DRM2 to add DNA methylation to nearby DNA. Both RdDM and the miRNA pathways involve related proteins from the Argonaute and Dicer families.[148]
Perhaps the most analogous pathways to RdDM in another eukaryotic kingdom are the sRNA directed transcriptional gene silencing (TGS) and co-transcriptional gene silencing (CTGS) pathways in Schizosaccharomyces pombe.[149] In S. pombe, TGS directs methylation of H3K9, leading to heterochromatin formation, and is directed by sRNAs produced from the targeted regions.[150] Similar to canonical RdDM, this pathway is a positive feedback loop: sRNAs are generated preferentially from heterochromatin-rich areas of the genome, and these sRNAs direct the addition of K3K9 methylation to maintain/spread heterochromatin. Meanwhile, CTGS is directed by AGO1-bound sRNAs, similar to PTGS within plants, and results in the inhibition of transcription by Pol II, as well as to Pol II release.[151][152] Unlike RdDM, TGS and CTGS in S. pombe do not rely on transcription from non-Pol II sources or lead to the addition of DNA methylation. However, the S. pombe pathways and RdDM share many of the same components, like RNA-directed RNA polymerases and sRNAs, and have similar functions in maintaining heterochromatin.
History
Introducing transgenes into organisms has been a widely used tool in plant genetics research for decades. However, researchers often find that their introduced transgenes are not expressed as strongly as expected, or sometimes even at all, a phenomenon called transgene silencing.[153] The discovery of transgene silencing in the 1990s spurred a great deal of interest in understanding the mechanisms behind this silencing.[154][155][156] Researchers found that transgene silencing was ubiquitous, occurring in multiple species (including Arabidopsis, Tobacco, and Petunia), and was associated with increased DNA methylation over and around the silenced transgene.[157][158][159]
Around the same time in 1994, work in tobacco plants had revealed a new pathway involving RNAs that resulted in DNA methylation. Researchers found that when viroids were introduced into the plant and integrated into the plant genome, the viroid sequences, but not the host genome, gained DNA methylation.[49] The deposition of methylation over these foreign viroid sequences helped inhibit viroid replication, and was therefore thought to represent a plant pathogen defense mechanism. The evidence suggested that the viroid RNAs produced during viroid replication were being used by the plant as a template to help target DNA methylation to the viroid sequences. This mechanism was therefore named RNA-directed DNA methylation, or RdDM.[49]
RdDM turned out to be the solution to the transgene mystery: like viroids and viruses, transgenes are foreign sequences, and as a result they are often recognized as foreign invaders and targeted for silencing by RdDM and PTGS. Since transgene silencing was a reliable marker of RdDM activity, researchers were able to design genetic screens to identify mutants that failed to trigger silencing at transgenes, reasoning that these genes were likely to be involved in the RdDM pathway. These experiments revealed many parts of the pathway, including RNA Pol IV and V, Dicer-like proteins, Argonautes, and others.[6][160][161]
The involvement of sRNAs in RdDM was initially suspected due to the similarity between RdDM and RNAi, the latter of which had recently been shown to involve small RNAs.[49][162] To test whether sRNAs were involved in RdDM, RNA hairpin structures complementary to a specific gene promoter were introduced into Arabidopsis and Tobacco.[163] The hairpin RNAs were processed into sRNAs, which were able to trigger the addition of DNA methylation to the targeted promoter and silence the gene.[163] This demonstrated that sRNAs could direct DNA methylation to specific loci. Later efforts showed that the sRNAs involved in RdDM were approximately 24-26 nt long, while the sRNAs associated with RNAi were only about 21-22 nt in length.[164][165] Soon after, the identification of AGO4 and characterization of its role in RdDM led to predictions, later confirmed, that 24 nt sRNAs were associating with AGO4 and directing DNA methylation to complementary loci.[166][165]
Early work on transgene silencing and RdDM also identified SDE4 as required for the production of most sRNAs involved in RdDM.[167] SDE4 would later be identified as the largest subunit of Pol IV, and renamed NRPD1. A number of studies published in quick succession from multiple research groups, utilizing both forward and reverse genetic approaches, went on to identify and characterize Pol IV and Pol V as highly specialized plant RNA polymerases involved in RdDM.[168][169][170][171] The Pol IV / Pol V naming convention was adopted shortly thereafter.[88][141]
Potential biotechnology applications
Since the mechanism underlying the sequence-specificity of RdDM is well known, RdDM can be ‘tricked’ into targeting and silencing endogenous genes in a highly specific manner, which has a number of potential biotechnological and bioengineering applications. Several different methods can be used to trigger RdDM-based DNA methylation and silencing of specific genes. One method, called virus-induced gene silencing (VIGS), involves inserting part of the promoter sequence of the desired target gene into a virus.[172] The virus will reproduce the chunk of promoter sequence as part of its own RNA, which is otherwise foreign to the plant. Because the viral RNA is foreign, it will be targeted for PTGS and processed into sRNAs, some of which will be complementary to the original target gene's promoter. A subset of these sRNAs will recruit the RdDM machinery to the target gene to add DNA methylation. In one study, researchers used this method with an engineered Cucumber Mosaic Virus to recruit RdDM to silence a gene that affected flower pigmentation in petunia, and another that affected fruit ripening in tomato.[173] In both cases, they showed that DNA methylation was added to the locus as expected. In petunia, both the gain of DNA methylation and changes in flower coloration were heritable, while only partial silencing and heritability were observed in tomato. VIGS has also been used to silence the FLOWERING WAGENINGEN (FWA) locus in Arabidopsis, which resulted in plants that flowered later than normal.[172] The same study also showed that the inhibitory effect of VIGS on FWA and flowering can become stronger over the course of successful generations.[172]
Another method to target RdDM to a desired target gene involves introducing a hairpin RNA construct that is complementary to the target locus. Hairpin RNAs contain an inverted repeat, which causes the RNA molecule to form a double-stranded RNA (dsRNA) structure called an RNA hairpin. The dsRNA hairpin can be processed by DCL proteins into sRNAs which are complementary to the target locus, triggering RdDM at that locus. This method has been used in several studies.[12][174][175]
Changes induced by RdDM can sometimes be maintained and inherited over multiple generations without outside intervention or manipulation, suggesting that RdDM can be a valuable tool for targeted epigenome editing. Recent work has even bypassed RdDM altogether by artificially tethering DRM2 (or other components of the RdDM pathway) directly to specific target loci, using either zinc finger nucleases or CRISPR.[90][176] In these experiments, tethering the RdDM machinery to a specific locus led to gain of DNA methylation at the target site that was often heritable for multiple generations, even once the artificial construct was removed through crossing. For all of these methods, however, more work on minimizing off-target effects and increasing DNA methylation efficiency is needed.
Genetically Modified Organisms (GMOs) have played a large role in recent agricultural research and practice, but have proven controversial, and face regulatory barriers to implementation in some jurisdictions. GMOs are defined by the inclusion of “foreign” genetic material into the genome. The treatment of plants with engineered RNAs or viruses intended to trigger RdDM does not change the underlying DNA sequence of the treated plant's genome; only the epigenetic state of portions of the DNA sequence already present are altered. As a result, these plants are not considered GMOs. This has led to efforts to utilize RdDM and other RNA-mediated effects to induce agriculturally-beneficial traits, like altering pathogen or herbicide susceptibility, or speeding up plant breeding by quickly inducing favorable traits.[177][178][179] However, while this is an area of active interest, there are few broadly implemented applications as of now.
References
![]() This article was adapted from the following source under a CC BY 4.0 license (2020) (reviewer reports):
Robert M. Erdmann; Colette Lafontaine Picard (8 October 2020). "RNA-directed DNA Methylation". PLOS Genetics. 16 (10): e1009034. doi:10.1371/JOURNAL.PGEN.1009034. ISSN 1553-7390. PMID 33031395. Wikidata Q100233435.
This article was adapted from the following source under a CC BY 4.0 license (2020) (reviewer reports):
Robert M. Erdmann; Colette Lafontaine Picard (8 October 2020). "RNA-directed DNA Methylation". PLOS Genetics. 16 (10): e1009034. doi:10.1371/JOURNAL.PGEN.1009034. ISSN 1553-7390. PMID 33031395. Wikidata Q100233435.{{cite journal}}: CS1 maint: unflagged free DOI (link)
- 1 2 3 4 Dubin MJ, Mittelsten Scheid O, Becker C (April 2018). "Transposons: a blessing curse". Current Opinion in Plant Biology. 42: 23–29. doi:10.1016/j.pbi.2018.01.003. PMID 29453028.
- ↑ Wicker T, Gundlach H, Spannagl M, Uauy C, Borrill P, Ramírez-González RH, et al. (August 2018). "Impact of transposable elements on genome structure and evolution in bread wheat". Genome Biology. 19 (1): 103. doi:10.1186/s13059-018-1479-0. PMC 6097303. PMID 30115100.
- 1 2 3 4 5 6 7 8 9 10 11 Sigman MJ, Slotkin RK (February 2016). "The First Rule of Plant Transposable Element Silencing: Location, Location, Location". The Plant Cell. 28 (2): 304–13. doi:10.1105/tpc.15.00869. PMC 4790875. PMID 26869697.
- ↑ Deniz Ö, Frost JM, Branco MR (July 2019). "Regulation of transposable elements by DNA modifications". Nature Reviews. Genetics. 20 (7): 417–431. doi:10.1038/s41576-019-0106-6. PMID 30867571. S2CID 76662244.
- 1 2 3 4 5 6 7 8 9 10 Zemach A, Kim MY, Hsieh PH, Coleman-Derr D, Eshed-Williams L, Thao K, et al. (March 2013). "The Arabidopsis nucleosome remodeler DDM1 allows DNA methyltransferases to access H1-containing heterochromatin". Cell. 153 (1): 193–205. doi:10.1016/j.cell.2013.02.033. PMC 4035305. PMID 23540698.
- 1 2 3 Chan SW, Zilberman D, Xie Z, Johansen LK, Carrington JC, Jacobsen SE (February 2004). "RNA silencing genes control de novo DNA methylation". Science. 303 (5662): 1336. doi:10.1126/science.1095989. PMID 14988555. S2CID 44659873.
- ↑ Pérez-Hormaeche J, Potet F, Beauclair L, Le Masson I, Courtial B, Bouché N, Lucas H (July 2008). "Invasion of the Arabidopsis genome by the tobacco retrotransposon Tnt1 is controlled by reversible transcriptional gene silencing". Plant Physiology. 147 (3): 1264–78. doi:10.1104/pp.108.117846. PMC 2442547. PMID 18467467.
- 1 2 Nuthikattu S, McCue AD, Panda K, Fultz D, DeFraia C, Thomas EN, Slotkin RK (May 2013). "The initiation of epigenetic silencing of active transposable elements is triggered by RDR6 and 21-22 nucleotide small interfering RNAs". Plant Physiology. 162 (1): 116–31. doi:10.1104/pp.113.216481. PMC 3641197. PMID 23542151.
- 1 2 3 4 5 6 Marí-Ordóñez A, Marchais A, Etcheverry M, Martin A, Colot V, Voinnet O (September 2013). "Reconstructing de novo silencing of an active plant retrotransposon". Nature Genetics. 45 (9): 1029–39. doi:10.1038/ng.2703. PMID 23852169. S2CID 13122409.
- 1 2 3 4 5 McCue AD, Panda K, Nuthikattu S, Choudury SG, Thomas EN, Slotkin RK (January 2015). "ARGONAUTE 6 bridges transposable element mRNA-derived siRNAs to the establishment of DNA methylation". The EMBO Journal. 34 (1): 20–35. doi:10.15252/embj.201489499. PMC 4291478. PMID 25388951.
- ↑ Harris CJ, Scheibe M, Wongpalee SP, Liu W, Cornett EM, Vaughan RM, et al. (December 2018). "A DNA methylation reader complex that enhances gene transcription". Science. 362 (6419): 1182–1186. Bibcode:2018Sci...362.1182H. doi:10.1126/science.aar7854. PMC 6353633. PMID 30523112.
- 1 2 3 4 5 6 Williams BP, Pignatta D, Henikoff S, Gehring M (March 2015). "Methylation-sensitive expression of a DNA demethylase gene serves as an epigenetic rheostat". PLOS Genetics. 11 (3): e1005142. doi:10.1371/journal.pgen.1005142. PMC 4380477. PMID 25826366.
- 1 2 3 4 Lei M, Zhang H, Julian R, Tang K, Xie S, Zhu JK (March 2015). "Regulatory link between DNA methylation and active demethylation in Arabidopsis". Proceedings of the National Academy of Sciences of the United States of America. 112 (11): 3553–7. Bibcode:2015PNAS..112.3553L. doi:10.1073/pnas.1502279112. PMC 4371987. PMID 25733903.
- 1 2 Penterman J, Zilberman D, Huh JH, Ballinger T, Henikoff S, Fischer RL (April 2007). "DNA demethylation in the Arabidopsis genome". Proceedings of the National Academy of Sciences of the United States of America. 104 (16): 6752–7. Bibcode:2007PNAS..104.6752P. doi:10.1073/pnas.0701861104. PMC 1847597. PMID 17409185.
- ↑ Cho J (2018). "Transposon-Derived Non-coding RNAs and Their Function in Plants". Frontiers in Plant Science. 9: 600. doi:10.3389/fpls.2018.00600. PMC 5943564. PMID 29774045.
- ↑ Mirouze M, Reinders J, Bucher E, Nishimura T, Schneeberger K, Ossowski S, et al. (September 2009). "Selective epigenetic control of retrotransposition in Arabidopsis". Nature. 461 (7262): 427–30. Bibcode:2009Natur.461..427M. doi:10.1038/nature08328. PMID 19734882. S2CID 205218044.
- 1 2 Ito H, Gaubert H, Bucher E, Mirouze M, Vaillant I, Paszkowski J (April 2011). "An siRNA pathway prevents transgenerational retrotransposition in plants subjected to stress". Nature. 472 (7341): 115–9. Bibcode:2011Natur.472..115I. doi:10.1038/nature09861. PMID 21399627. S2CID 4426724.
- 1 2 Cavrak VV, Lettner N, Jamge S, Kosarewicz A, Bayer LM, Mittelsten Scheid O (January 2014). "How a retrotransposon exploits the plant's heat stress response for its activation". PLOS Genetics. 10 (1): e1004115. doi:10.1371/journal.pgen.1004115. PMC 3907296. PMID 24497839.
- 1 2 Soppe WJ, Jacobsen SE, Alonso-Blanco C, Jackson JP, Kakutani T, Koornneef M, Peeters AJ (October 2000). "The late flowering phenotype of fwa mutants is caused by gain-of-function epigenetic alleles of a homeodomain gene". Molecular Cell. 6 (4): 791–802. doi:10.1016/s1097-2765(05)00090-0. PMID 11090618.
- 1 2 Kinoshita Y, Saze H, Kinoshita T, Miura A, Soppe WJ, Koornneef M, Kakutani T (January 2007). "Control of FWA gene silencing in Arabidopsis thaliana by SINE-related direct repeats". The Plant Journal. 49 (1): 38–45. doi:10.1111/j.1365-313X.2006.02936.x. hdl:11858/00-001M-0000-0012-38D2-5. PMID 17144899.
- ↑ Gouil Q, Baulcombe DC (December 2016). "DNA Methylation Signatures of the Plant Chromomethyltransferases". PLOS Genetics. 12 (12): e1006526. doi:10.1371/journal.pgen.1006526. PMC 5221884. PMID 27997534.
- ↑ Grover JW, Kendall T, Baten A, Burgess D, Freeling M, King GJ, Mosher RA (May 2018). "Maternal components of RNA-directed DNA methylation are required for seed development in Brassica rapa". The Plant Journal. 94 (4): 575–582. doi:10.1111/tpj.13910. hdl:10150/628261. PMID 29569777. S2CID 4212729.
- 1 2 Wang G, Köhler C (February 2017). "Epigenetic processes in flowering plant reproduction". Journal of Experimental Botany. 68 (4): 797–807. doi:10.1093/jxb/erw486. PMID 28062591. S2CID 23237961.
- 1 2 3 Martinez G, Köhler C (April 2017). "Role of small RNAs in epigenetic reprogramming during plant sexual reproduction". Current Opinion in Plant Biology. 36: 22–28. doi:10.1016/j.pbi.2016.12.006. PMID 28088028.
- 1 2 Olmedo-Monfil V, Durán-Figueroa N, Arteaga-Vázquez M, Demesa-Arévalo E, Autran D, Grimanelli D, et al. (March 2010). "Control of female gamete formation by a small RNA pathway in Arabidopsis". Nature. 464 (7288): 628–32. Bibcode:2010Natur.464..628O. doi:10.1038/nature08828. PMC 4613780. PMID 20208518.
- ↑ Slotkin RK, Vaughn M, Borges F, Tanurdzić M, Becker JD, Feijó JA, Martienssen RA (February 2009). "Epigenetic reprogramming and small RNA silencing of transposable elements in pollen". Cell. 136 (3): 461–72. doi:10.1016/j.cell.2008.12.038. PMC 2661848. PMID 19203581.
- 1 2 Martínez G, Panda K, Köhler C, Slotkin RK (March 2016). "Silencing in sperm cells is directed by RNA movement from the surrounding nurse cell". Nature Plants. 2 (4): 16030. doi:10.1038/nplants.2016.30. PMID 27249563. S2CID 24746649.
- ↑ Erdmann RM, Hoffmann A, Walter HK, Wagenknecht HA, Groß-Hardt R, Gehring M (September 2017). "Molecular movement in the Arabidopsis thaliana female gametophyte". Plant Reproduction. 30 (3): 141–146. doi:10.1007/s00497-017-0304-3. PMC 5599461. PMID 28695277.
- 1 2 3 Siomi MC, Sato K, Pezic D, Aravin AA (April 2011). "PIWI-interacting small RNAs: the vanguard of genome defence". Nature Reviews. Molecular Cell Biology. 12 (4): 246–58. doi:10.1038/nrm3089. PMID 21427766. S2CID 5710813.
- 1 2 3 Ernst C, Odom DT, Kutter C (November 2017). "The emergence of piRNAs against transposon invasion to preserve mammalian genome integrity". Nature Communications. 8 (1): 1411. Bibcode:2017NatCo...8.1411E. doi:10.1038/s41467-017-01049-7. PMC 5681665. PMID 29127279.
- 1 2 Kawakatsu T, Stuart T, Valdes M, Breakfield N, Schmitz RJ, Nery JR, et al. (April 2016). "Unique cell-type-specific patterns of DNA methylation in the root meristem". Nature Plants. 2 (5): 16058. doi:10.1038/nplants.2016.58. PMC 4855458. PMID 27243651.
- ↑ Vu TM, Nakamura M, Calarco JP, Susaki D, Lim PQ, Kinoshita T, et al. (July 2013). "RNA-directed DNA methylation regulates parental genomic imprinting at several loci in Arabidopsis". Development. 140 (14): 2953–60. doi:10.1242/dev.092981. PMC 3879202. PMID 23760956.
- ↑ Waters AJ, Bilinski P, Eichten SR, Vaughn MW, Ross-Ibarra J, Gehring M, Springer NM (November 2013). "Comprehensive analysis of imprinted genes in maize reveals allelic variation for imprinting and limited conservation with other species". Proceedings of the National Academy of Sciences of the United States of America. 110 (48): 19639–44. Bibcode:2013PNAS..11019639W. doi:10.1073/pnas.1309182110. PMC 3845156. PMID 24218619.
- ↑ Pignatta D, Erdmann RM, Scheer E, Picard CL, Bell GW, Gehring M (July 2014). "Natural epigenetic polymorphisms lead to intraspecific variation in Arabidopsis gene imprinting". eLife. 3: e03198. doi:10.7554/eLife.03198. PMC 4115658. PMID 24994762.
- ↑ Klosinska M, Picard CL, Gehring M (September 2016). "Conserved imprinting associated with unique epigenetic signatures in the Arabidopsis genus". Nature Plants. 2 (10): 16145. doi:10.1038/nplants.2016.145. PMC 5367468. PMID 27643534.
- ↑ Hatorangan MR, Laenen B, Steige KA, Slotte T, Köhler C (August 2016). "Rapid Evolution of Genomic Imprinting in Two Species of the Brassicaceae". The Plant Cell. 28 (8): 1815–27. doi:10.1105/tpc.16.00304. PMC 5006707. PMID 27465027.
- ↑ Erdmann RM, Satyaki PR, Klosinska M, Gehring M (December 2017). "A Small RNA Pathway Mediates Allelic Dosage in Endosperm". Cell Reports. 21 (12): 3364–3372. doi:10.1016/j.celrep.2017.11.078. PMID 29262317.
- ↑ Satyaki PR, Gehring M (July 2019). "Paternally Acting Canonical RNA-Directed DNA Methylation Pathway Genes Sensitize Arabidopsis Endosperm to Paternal Genome Dosage". The Plant Cell. 31 (7): 1563–1578. doi:10.1105/tpc.19.00047. PMC 6635864. PMID 31064867.
- ↑ Iwasaki M, Hyvärinen L, Piskurewicz U, Lopez-Molina L (March 2019). "Non-canonical RNA-directed DNA methylation participates in maternal and environmental control of seed dormancy". eLife. 8. doi:10.7554/eLife.37434. PMC 6435323. PMID 30910007.
- ↑ Cheng J, Niu Q, Zhang B, Chen K, Yang R, Zhu JK, et al. (December 2018). "Downregulation of RdDM during strawberry fruit ripening". Genome Biology. 19 (1): 212. doi:10.1186/s13059-018-1587-x. PMC 6280534. PMID 30514401.
- ↑ Guo X, Ma Z, Zhang Z, Cheng L, Zhang X, Li T (2017). "Small RNA-Sequencing Links Physiological Changes and RdDM Process to Vegetative-to-Floral Transition in Apple". Frontiers in Plant Science. 8: 873. doi:10.3389/fpls.2017.00873. PMC 5447065. PMID 28611800.
- ↑ Fortes AM, Gallusci P (2017-02-06). "Plant Stress Responses and Phenotypic Plasticity in the Epigenomics Era: Perspectives on the Grapevine Scenario, a Model for Perennial Crop Plants". Frontiers in Plant Science. 8: 82. doi:10.3389/fpls.2017.00082. PMC 5292615. PMID 28220131.
- ↑ Kumar A, Bennetzen JL (1999). "Plant retrotransposons". Annual Review of Genetics. 33: 479–532. doi:10.1146/annurev.genet.33.1.479. PMID 10690416.
- ↑ Ito H, Kim JM, Matsunaga W, Saze H, Matsui A, Endo TA, et al. (March 2016). "A Stress-Activated Transposon in Arabidopsis Induces Transgenerational Abscisic Acid Insensitivity". Scientific Reports. 6 (1): 23181. Bibcode:2016NatSR...623181I. doi:10.1038/srep23181. PMC 4791638. PMID 26976262.
- ↑ Liu J, Feng L, Li J, He Z (2015-04-24). "Genetic and epigenetic control of plant heat responses". Frontiers in Plant Science. 6: 267. doi:10.3389/fpls.2015.00267. PMC 4408840. PMID 25964789.
- ↑ Popova OV, Dinh HQ, Aufsatz W, Jonak C (March 2013). "The RdDM pathway is required for basal heat tolerance in Arabidopsis". Molecular Plant. 6 (2): 396–410. doi:10.1093/mp/sst023. PMC 3603006. PMID 23376771.
- ↑ Tricker PJ, Gibbings JG, Rodríguez López CM, Hadley P, Wilkinson MJ (June 2012). "Low relative humidity triggers RNA-directed de novo DNA methylation and suppression of genes controlling stomatal development". Journal of Experimental Botany. 63 (10): 3799–813. doi:10.1093/jxb/ers076. PMC 3733579. PMID 22442411.
- ↑ Xu R, Wang Y, Zheng H, Lu W, Wu C, Huang J, et al. (September 2015). "Salt-induced transcription factor MYB74 is regulated by the RNA-directed DNA methylation pathway in Arabidopsis". Journal of Experimental Botany. 66 (19): 5997–6008. doi:10.1093/jxb/erv312. PMC 4566987. PMID 26139822.
- 1 2 3 4 Wassenegger M, Heimes S, Riedel L, Sänger HL (February 1994). "RNA-directed de novo methylation of genomic sequences in plants". Cell. 76 (3): 567–76. doi:10.1016/0092-8674(94)90119-8. PMID 8313476. S2CID 35858018.
- 1 2 3 4 Huang J, Yang M, Zhang X (April 2016). "The function of small RNAs in plant biotic stress response". Journal of Integrative Plant Biology. 58 (4): 312–27. doi:10.1111/jipb.12463. PMID 26748943.
- ↑ Raja P, Jackel JN, Li S, Heard IM, Bisaro DM (March 2014). "Arabidopsis double-stranded RNA binding protein DRB3 participates in methylation-mediated defense against geminiviruses". Journal of Virology. 88 (5): 2611–22. doi:10.1128/JVI.02305-13. PMC 3958096. PMID 24352449.
- ↑ Jackel JN, Storer JM, Coursey T, Bisaro DM (August 2016). Simon A (ed.). "Arabidopsis RNA Polymerases IV and V Are Required To Establish H3K9 Methylation, but Not Cytosine Methylation, on Geminivirus Chromatin". Journal of Virology. 90 (16): 7529–7540. doi:10.1128/JVI.00656-16. PMC 4984644. PMID 27279611.
- ↑ Calil IP, Fontes EP (March 2017). "Plant immunity against viruses: antiviral immune receptors in focus". Annals of Botany. 119 (5): 711–723. doi:10.1093/aob/mcw200. PMC 5604577. PMID 27780814.
- 1 2 3 4 5 6 7 8 9 Matzke MA, Mosher RA (June 2014). "RNA-directed DNA methylation: an epigenetic pathway of increasing complexity". Nature Reviews. Genetics. 15 (6): 394–408. doi:10.1038/nrg3683. PMID 24805120. S2CID 54489227.
- ↑ Wang MB, Masuta C, Smith NA, Shimura H (October 2012). "RNA silencing and plant viral diseases". Molecular Plant-Microbe Interactions. 25 (10): 1275–85. doi:10.1094/MPMI-04-12-0093-CR. PMID 22670757.
- ↑ Wang Y, Wu Y, Gong Q, Ismayil A, Yuan Y, Lian B, et al. (March 2019). Simon AE (ed.). "Geminiviral V2 Protein Suppresses Transcriptional Gene Silencing through Interaction with AGO4". Journal of Virology. 93 (6): e01675–18, /jvi/93/6/JVI.01675–18.atom. doi:10.1128/JVI.01675-18. PMC 6401443. PMID 30626668.
- 1 2 Dowen RH, Pelizzola M, Schmitz RJ, Lister R, Dowen JM, Nery JR, et al. (August 2012). "Widespread dynamic DNA methylation in response to biotic stress". Proceedings of the National Academy of Sciences of the United States of America. 109 (32): E2183-91. doi:10.1073/pnas.1209329109. PMC 3420206. PMID 22733782.
- ↑ López A, Ramírez V, García-Andrade J, Flors V, Vera P (December 2011). Pikaard CS (ed.). "The RNA silencing enzyme RNA polymerase v is required for plant immunity". PLOS Genetics. 7 (12): e1002434. doi:10.1371/journal.pgen.1002434. PMC 3248562. PMID 22242006.
- 1 2 Rasmann S, De Vos M, Casteel CL, Tian D, Halitschke R, Sun JY, et al. (February 2012). "Herbivory in the previous generation primes plants for enhanced insect resistance". Plant Physiology. 158 (2): 854–63. doi:10.1104/pp.111.187831. PMC 3271773. PMID 22209873.
- ↑ Gohlke J, Scholz CJ, Kneitz S, Weber D, Fuchs J, Hedrich R, Deeken R (2013-02-07). McDowell JM (ed.). "DNA methylation mediated control of gene expression is critical for development of crown gall tumors". PLOS Genetics. 9 (2): e1003267. doi:10.1371/journal.pgen.1003267. PMC 3567176. PMID 23408907.
- ↑ Espinas NA, Saze H, Saijo Y (2016-08-11). "Epigenetic Control of Defense Signaling and Priming in Plants". Frontiers in Plant Science. 7: 1201. doi:10.3389/fpls.2016.01201. PMC 4980392. PMID 27563304.
- ↑ Aufsatz W, Mette MF, van der Winden J, Matzke AJ, Matzke M (December 2002). "RNA-directed DNA methylation in Arabidopsis". Proceedings of the National Academy of Sciences of the United States of America. 99 Suppl 4 (Supplement 4): 16499–506. Bibcode:2002PNAS...9916499A. doi:10.1073/pnas.162371499. PMC 139914. PMID 12169664.
- 1 2 Matzke MA, Primig M, Trnovsky J, Matzke AJ (March 1989). "Reversible methylation and inactivation of marker genes in sequentially transformed tobacco plants". The EMBO Journal. 8 (3): 643–9. doi:10.1002/j.1460-2075.1989.tb03421.x. PMC 400855. PMID 16453872.
- ↑ Gutzat R, Mittelsten Scheid O (November 2012). "Epigenetic responses to stress: triple defense?". Current Opinion in Plant Biology. 15 (5): 568–73. doi:10.1016/j.pbi.2012.08.007. PMC 3508409. PMID 22960026.
- ↑ Boyko A, Kovalchuk I (August 2010). Shiu SH (ed.). "Transgenerational response to stress in Arabidopsis thaliana". Plant Signaling & Behavior. 5 (8): 995–8. Bibcode:2010PlSiB...5..995B. doi:10.4161/psb.5.8.12227. PMC 3115178. PMID 20724818.
- ↑ Mermigka G, Verret F, Kalantidis K (April 2016). "RNA silencing movement in plants". Journal of Integrative Plant Biology. 58 (4): 328–42. doi:10.1111/jipb.12423. PMID 26297506.
- ↑ Lewsey MG, Hardcastle TJ, Melnyk CW, Molnar A, Valli A, Urich MA, et al. (February 2016). "Mobile small RNAs regulate genome-wide DNA methylation". Proceedings of the National Academy of Sciences of the United States of America. 113 (6): E801-10. Bibcode:2016PNAS..113E.801L. doi:10.1073/pnas.1515072113. PMC 4760824. PMID 26787884.
- 1 2 Tamiru M, Hardcastle TJ, Lewsey MG (January 2018). "Regulation of genome-wide DNA methylation by mobile small RNAs". The New Phytologist. 217 (2): 540–546. doi:10.1111/nph.14874. PMID 29105762.
- 1 2 3 Molnar A, Melnyk CW, Bassett A, Hardcastle TJ, Dunn R, Baulcombe DC (May 2010). "Small silencing RNAs in plants are mobile and direct epigenetic modification in recipient cells". Science. 328 (5980): 872–5. Bibcode:2010Sci...328..872M. doi:10.1126/science.1187959. PMID 20413459. S2CID 206525853.
- ↑ Bai S, Kasai A, Yamada K, Li T, Harada T (August 2011). "A mobile signal transported over a long distance induces systemic transcriptional gene silencing in a grafted partner". Journal of Experimental Botany. 62 (13): 4561–70. doi:10.1093/jxb/err163. PMC 3170550. PMID 21652532.
- ↑ Zhang W, Kollwig G, Stecyk E, Apelt F, Dirks R, Kragler F (October 2014). "Graft-transmissible movement of inverted-repeat-induced siRNA signals into flowers". The Plant Journal. 80 (1): 106–21. doi:10.1111/tpj.12622. PMID 25039964.
- ↑ Parent JS, Martínez de Alba AE, Vaucheret H (2012). "The origin and effect of small RNA signaling in plants". Frontiers in Plant Science. 3: 179. doi:10.3389/fpls.2012.00179. PMC 3414853. PMID 22908024.
- 1 2 Stroud H, Do T, Du J, Zhong X, Feng S, Johnson L, et al. (January 2014). "Non-CG methylation patterns shape the epigenetic landscape in Arabidopsis". Nature Structural & Molecular Biology. 21 (1): 64–72. doi:10.1038/nsmb.2735. PMC 4103798. PMID 24336224.
- 1 2 Bewick AJ, Niederhuth CE, Ji L, Rohr NA, Griffin PT, Leebens-Mack J, Schmitz RJ (May 2017). "The evolution of CHROMOMETHYLASES and gene body DNA methylation in plants". Genome Biology. 18 (1): 65. doi:10.1186/s13059-017-1195-1. PMC 5410703. PMID 28457232.
- ↑ Bartels A, Han Q, Nair P, Stacey L, Gaynier H, Mosley M, et al. (July 2018). "Dynamic DNA Methylation in Plant Growth and Development". International Journal of Molecular Sciences. 19 (7): 2144. doi:10.3390/ijms19072144. PMC 6073778. PMID 30041459.
- ↑ Wendte JM, Schmitz RJ (March 2018). "Specifications of Targeting Heterochromatin Modifications in Plants". Molecular Plant. 11 (3): 381–387. doi:10.1016/j.molp.2017.10.002. PMID 29032247.
- ↑ Law JA, Jacobsen SE (March 2010). "Establishing, maintaining and modifying DNA methylation patterns in plants and animals". Nature Reviews. Genetics. 11 (3): 204–20. doi:10.1038/nrg2719. PMC 3034103. PMID 20142834.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 Cuerda-Gil D, Slotkin RK (November 2016). "Non-canonical RNA-directed DNA methylation". Nature Plants. 2 (11): 16163. doi:10.1038/nplants.2016.163. PMID 27808230. S2CID 4248951.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 Matzke MA, Kanno T, Matzke AJ (2015). "RNA-Directed DNA Methylation: The Evolution of a Complex Epigenetic Pathway in Flowering Plants". Annual Review of Plant Biology. 66: 243–67. doi:10.1146/annurev-arplant-043014-114633. PMID 25494460.
- 1 2 3 4 Wendte JM, Pikaard CS (January 2017). "The RNAs of RNA-directed DNA methylation". Biochimica et Biophysica Acta (BBA) - Gene Regulatory Mechanisms. 1860 (1): 140–148. doi:10.1016/j.bbagrm.2016.08.004. PMC 5203809. PMID 27521981.
- ↑ Zhai J, Bischof S, Wang H, Feng S, Lee TF, Teng C, et al. (October 2015). "A One Precursor One siRNA Model for Pol IV-Dependent siRNA Biogenesis". Cell. 163 (2): 445–55. doi:10.1016/j.cell.2015.09.032. PMC 5023148. PMID 26451488.
- 1 2 3 Blevins T, Podicheti R, Mishra V, Marasco M, Wang J, Rusch D, et al. (October 2015). "Identification of Pol IV and RDR2-dependent precursors of 24 nt siRNAs guiding de novo DNA methylation in Arabidopsis". eLife. 4: e09591. doi:10.7554/eLife.09591. PMC 4716838. PMID 26430765.
- 1 2 Singh J, Mishra V, Wang F, Huang HY, Pikaard CS (August 2019). "Reaction Mechanisms of Pol IV, RDR2, and DCL3 Drive RNA Channeling in the siRNA-Directed DNA Methylation Pathway". Molecular Cell. 75 (3): 576–589.e5. doi:10.1016/j.molcel.2019.07.008. PMC 6698059. PMID 31398324.
- ↑ Panda K, Ji L, Neumann DA, Daron J, Schmitz RJ, Slotkin RK (August 2016). "Full-length autonomous transposable elements are preferentially targeted by expression-dependent forms of RNA-directed DNA methylation". Genome Biology. 17 (1): 170. doi:10.1186/s13059-016-1032-y. PMC 4977677. PMID 27506905.
- ↑ Zhang Z, Liu X, Guo X, Wang XJ, Zhang X (April 2016). "Arabidopsis AGO3 predominantly recruits 24-nt small RNAs to regulate epigenetic silencing". Nature Plants. 2 (5): 16049. doi:10.1038/nplants.2016.49. PMID 27243648. S2CID 8933827.
- ↑ Meister G (July 2013). "Argonaute proteins: functional insights and emerging roles". Nature Reviews. Genetics. 14 (7): 447–59. doi:10.1038/nrg3462. PMID 23732335. S2CID 5210500.
- 1 2 3 Wierzbicki AT, Haag JR, Pikaard CS (November 2008). "Noncoding transcription by RNA polymerase Pol IVb/Pol V mediates transcriptional silencing of overlapping and adjacent genes". Cell. 135 (4): 635–48. doi:10.1016/j.cell.2008.09.035. PMC 2602798. PMID 19013275.
- ↑ Cao X, Jacobsen SE (July 2002). "Role of the arabidopsis DRM methyltransferases in de novo DNA methylation and gene silencing". Current Biology. 12 (13): 1138–44. doi:10.1016/s0960-9822(02)00925-9. PMID 12121623. S2CID 15695949.
- 1 2 3 Gallego-Bartolomé J, Liu W, Kuo PH, Feng S, Ghoshal B, Gardiner J, et al. (February 2019). "Co-targeting RNA Polymerases IV and V Promotes Efficient De Novo DNA Methylation in Arabidopsis". Cell. 176 (5): 1068–1082.e19. doi:10.1016/j.cell.2019.01.029. PMC 6386582. PMID 30739798.
- 1 2 Voinnet O (July 2008). "Use, tolerance and avoidance of amplified RNA silencing by plants". Trends in Plant Science. 13 (7): 317–28. doi:10.1016/j.tplants.2008.05.004. PMID 18565786.
- 1 2 Pontier D, Picart C, Roudier F, Garcia D, Lahmy S, Azevedo J, et al. (October 2012). "NERD, a plant-specific GW protein, defines an additional RNAi-dependent chromatin-based pathway in Arabidopsis". Molecular Cell. 48 (1): 121–32. doi:10.1016/j.molcel.2012.07.027. PMID 22940247.
- 1 2 3 4 Haag JR, Pikaard CS (July 2011). "Multisubunit RNA polymerases IV and V: purveyors of non-coding RNA for plant gene silencing". Nature Reviews. Molecular Cell Biology. 12 (8): 483–92. doi:10.1038/nrm3152. PMID 21779025. S2CID 9970159.
- 1 2 3 Zhou M, Law JA (October 2015). "RNA Pol IV and V in gene silencing: Rebel polymerases evolving away from Pol II's rules". Current Opinion in Plant Biology. 27: 154–64. doi:10.1016/j.pbi.2015.07.005. PMC 4618083. PMID 26344361.
- 1 2 Lahmy S, Pontier D, Bies-Etheve N, Laudié M, Feng S, Jobet E, et al. (December 2016). "Evidence for ARGONAUTE4-DNA interactions in RNA-directed DNA methylation in plants". Genes & Development. 30 (23): 2565–2570. doi:10.1101/gad.289553.116. PMC 5204349. PMID 27986858.
- 1 2 3 4 Henderson IR, Zhang X, Lu C, Johnson L, Meyers BC, Green PJ, Jacobsen SE (June 2006). "Dissecting Arabidopsis thaliana DICER function in small RNA processing, gene silencing and DNA methylation patterning". Nature Genetics. 38 (6): 721–5. doi:10.1038/ng1804. PMID 16699516. S2CID 10261689.
- 1 2 3 4 5 Bologna NG, Voinnet O (2014). "The diversity, biogenesis, and activities of endogenous silencing small RNAs in Arabidopsis". Annual Review of Plant Biology. 65: 473–503. doi:10.1146/annurev-arplant-050213-035728. PMID 24579988.
- ↑ Wang J, Mei J, Ren G (2019). "Plant microRNAs: Biogenesis, Homeostasis, and Degradation". Frontiers in Plant Science. 10: 360. doi:10.3389/fpls.2019.00360. PMC 6445950. PMID 30972093.
- 1 2 3 4 5 6 7 8 9 10 Stroud H, Greenberg MV, Feng S, Bernatavichute YV, Jacobsen SE (January 2013). "Comprehensive analysis of silencing mutants reveals complex regulation of the Arabidopsis methylome". Cell. 152 (1–2): 352–64. doi:10.1016/j.cell.2012.10.054. PMC 3597350. PMID 23313553.
- 1 2 3 4 Fang X, Qi Y (February 2016). "RNAi in Plants: An Argonaute-Centered View". The Plant Cell. 28 (2): 272–85. doi:10.1105/tpc.15.00920. PMC 4790879. PMID 26869699.
- ↑ Eun C, Lorkovic ZJ, Naumann U, Long Q, Havecker ER, Simon SA, et al. (2011). "AGO6 functions in RNA-mediated transcriptional gene silencing in shoot and root meristems in Arabidopsis thaliana". PLOS ONE. 6 (10): e25730. Bibcode:2011PLoSO...625730E. doi:10.1371/journal.pone.0025730. PMC 3187791. PMID 21998686.
- ↑ Durán-Figueroa N, Vielle-Calzada JP (November 2010). "ARGONAUTE9-dependent silencing of transposable elements in pericentromeric regions of Arabidopsis". Plant Signaling & Behavior. 5 (11): 1476–9. Bibcode:2010PlSiB...5.1476D. doi:10.4161/psb.5.11.13548. PMC 3115260. PMID 21057207.
- ↑ Cao X, Aufsatz W, Zilberman D, Mette MF, Huang MS, Matzke M, Jacobsen SE (December 2003). "Role of the DRM and CMT3 methyltransferases in RNA-directed DNA methylation". Current Biology. 13 (24): 2212–7. doi:10.1016/j.cub.2003.11.052. PMID 14680640. S2CID 8232599.
- ↑ Law JA, Vashisht AA, Wohlschlegel JA, Jacobsen SE (July 2011). "SHH1, a homeodomain protein required for DNA methylation, as well as RDR2, RDM4, and chromatin remodeling factors, associate with RNA polymerase IV". PLOS Genetics. 7 (7): e1002195. doi:10.1371/journal.pgen.1002195. PMC 3141008. PMID 21811420.
- ↑ Zhang H, Ma ZY, Zeng L, Tanaka K, Zhang CJ, Ma J, et al. (May 2013). "DTF1 is a core component of RNA-directed DNA methylation and may assist in the recruitment of Pol IV". Proceedings of the National Academy of Sciences of the United States of America. 110 (20): 8290–5. Bibcode:2013PNAS..110.8290Z. doi:10.1073/pnas.1300585110. PMC 3657815. PMID 23637343.
- 1 2 Law JA, Du J, Hale CJ, Feng S, Krajewski K, Palanca AM, et al. (June 2013). "Polymerase IV occupancy at RNA-directed DNA methylation sites requires SHH1". Nature. 498 (7454): 385–9. Bibcode:2013Natur.498..385L. doi:10.1038/nature12178. PMC 4119789. PMID 23636332.
- 1 2 3 Zhou M, Palanca AM, Law JA (June 2018). "Locus-specific control of the de novo DNA methylation pathway in Arabidopsis by the CLASSY family". Nature Genetics. 50 (6): 865–873. doi:10.1038/s41588-018-0115-y. PMC 6317521. PMID 29736015.
- 1 2 Yang DL, Zhang G, Wang L, Li J, Xu D, Di C, et al. (2018). "Four putative SWI2/SNF2 chromatin remodelers have dual roles in regulating DNA methylation in Arabidopsis". Cell Discovery. 4: 55. doi:10.1038/s41421-018-0056-8. PMC 6189096. PMID 30345072.
- ↑ Ji L, Chen X (April 2012). "Regulation of small RNA stability: methylation and beyond". Cell Research. 22 (4): 624–36. doi:10.1038/cr.2012.36. PMC 3317568. PMID 22410795.
- 1 2 3 4 Liu ZW, Shao CR, Zhang CJ, Zhou JX, Zhang SW, Li L, et al. (January 2014). "The SET domain proteins SUVH2 and SUVH9 are required for Pol V occupancy at RNA-directed DNA methylation loci". PLOS Genetics. 10 (1): e1003948. doi:10.1371/journal.pgen.1003948. PMC 3898904. PMID 24465213.
- ↑ Wierzbicki AT, Ream TS, Haag JR, Pikaard CS (May 2009). "RNA polymerase V transcription guides ARGONAUTE4 to chromatin". Nature Genetics. 41 (5): 630–4. doi:10.1038/ng.365. PMC 2674513. PMID 19377477.
- ↑ Zhong X, Hale CJ, Law JA, Johnson LM, Feng S, Tu A, Jacobsen SE (September 2012). "DDR complex facilitates global association of RNA polymerase V to promoters and evolutionarily young transposons". Nature Structural & Molecular Biology. 19 (9): 870–5. doi:10.1038/nsmb.2354. PMC 3443314. PMID 22864289.
- ↑ Pikaard CS, Haag JR, Pontes OM, Blevins T, Cocklin R (2012). "A transcription fork model for Pol IV and Pol V-dependent RNA-directed DNA methylation". Cold Spring Harbor Symposia on Quantitative Biology. 77: 205–12. doi:10.1101/sqb.2013.77.014803. PMID 23567894.
- ↑ He XJ, Hsu YF, Zhu S, Wierzbicki AT, Pontes O, Pikaard CS, et al. (May 2009). "An effector of RNA-directed DNA methylation in arabidopsis is an ARGONAUTE 4- and RNA-binding protein". Cell. 137 (3): 498–508. doi:10.1016/j.cell.2009.04.028. PMC 2700824. PMID 19410546.
- ↑ Liu W, Duttke SH, Hetzel J, Groth M, Feng S, Gallego-Bartolome J, et al. (March 2018). "RNA-directed DNA methylation involves co-transcriptional small-RNA-guided slicing of polymerase V transcripts in Arabidopsis". Nature Plants. 4 (3): 181–188. doi:10.1038/s41477-017-0100-y. PMC 5832601. PMID 29379150.
- 1 2 Zhu Y, Rowley MJ, Böhmdorfer G, Wierzbicki AT (January 2013). "A SWI/SNF chromatin-remodeling complex acts in noncoding RNA-mediated transcriptional silencing". Molecular Cell. 49 (2): 298–309. doi:10.1016/j.molcel.2012.11.011. PMC 3560041. PMID 23246435.
- ↑ Ausin I, Mockler TC, Chory J, Jacobsen SE (December 2009). "IDN1 and IDN2 are required for de novo DNA methylation in Arabidopsis thaliana". Nature Structural & Molecular Biology. 16 (12): 1325–7. doi:10.1038/nsmb.1690. PMC 2842998. PMID 19915591.
- ↑ Xie M, Ren G, Zhang C, Yu B (November 2012). "The DNA- and RNA-binding protein FACTOR of DNA METHYLATION 1 requires XH domain-mediated complex formation for its function in RNA-directed DNA methylation". The Plant Journal. 72 (3): 491–500. doi:10.1111/j.1365-313X.2012.05092.x. PMID 22757778.
- ↑ Jullien PE, Susaki D, Yelagandula R, Higashiyama T, Berger F (October 2012). "DNA methylation dynamics during sexual reproduction in Arabidopsis thaliana". Current Biology. 22 (19): 1825–30. doi:10.1016/j.cub.2012.07.061. PMID 22940470. S2CID 18586419.
- 1 2 3 4 Blevins T, Pontvianne F, Cocklin R, Podicheti R, Chandrasekhara C, Yerneni S, et al. (April 2014). "A two-step process for epigenetic inheritance in Arabidopsis". Molecular Cell. 54 (1): 30–42. doi:10.1016/j.molcel.2014.02.019. PMC 3988221. PMID 24657166.
- ↑ Peters AH, Kubicek S, Mechtler K, O'Sullivan RJ, Derijck AA, Perez-Burgos L, et al. (December 2003). "Partitioning and plasticity of repressive histone methylation states in mammalian chromatin". Molecular Cell. 12 (6): 1577–89. doi:10.1016/s1097-2765(03)00477-5. PMID 14690609.
- ↑ Jackson JP, Johnson L, Jasencakova Z, Zhang X, PerezBurgos L, Singh PB, et al. (March 2004). "Dimethylation of histone H3 lysine 9 is a critical mark for DNA methylation and gene silencing in Arabidopsis thaliana". Chromosoma. 112 (6): 308–15. doi:10.1007/s00412-004-0275-7. PMID 15014946. S2CID 17798608.
- 1 2 3 Du J, Johnson LM, Jacobsen SE, Patel DJ (September 2015). "DNA methylation pathways and their crosstalk with histone methylation". Nature Reviews. Molecular Cell Biology. 16 (9): 519–32. doi:10.1038/nrm4043. PMC 4672940. PMID 26296162.
- ↑ Li X, Harris CJ, Zhong Z, Chen W, Liu R, Jia B, et al. (September 2018). "Mechanistic insights into plant SUVH family H3K9 methyltransferases and their binding to context-biased non-CG DNA methylation". Proceedings of the National Academy of Sciences of the United States of America. 115 (37): E8793–E8802. Bibcode:2018PNAS..115E8793L. doi:10.1073/pnas.1809841115. PMC 6140468. PMID 30150382.
- ↑ Du J, Zhong X, Bernatavichute YV, Stroud H, Feng S, Caro E, et al. (September 2012). "Dual binding of chromomethylase domains to H3K9me2-containing nucleosomes directs DNA methylation in plants". Cell. 151 (1): 167–80. doi:10.1016/j.cell.2012.07.034. PMC 3471781. PMID 23021223.
- 1 2 Lachner M, O'Carroll D, Rea S, Mechtler K, Jenuwein T (March 2001). "Methylation of histone H3 lysine 9 creates a binding site for HP1 proteins". Nature. 410 (6824): 116–20. Bibcode:2001Natur.410..116L. doi:10.1038/35065132. PMID 11242053. S2CID 4331863.
- ↑ Mylne JS, Barrett L, Tessadori F, Mesnage S, Johnson L, Bernatavichute YV, et al. (March 2006). "LHP1, the Arabidopsis homologue of HETEROCHROMATIN PROTEIN1, is required for epigenetic silencing of FLC". Proceedings of the National Academy of Sciences of the United States of America. 103 (13): 5012–7. Bibcode:2006PNAS..103.5012M. doi:10.1073/pnas.0507427103. PMC 1458786. PMID 16549797.
- ↑ Zhao S, Cheng L, Gao Y, Zhang B, Zheng X, Wang L, et al. (January 2019). "Plant HP1 protein ADCP1 links multivalent H3K9 methylation readout to heterochromatin formation". Cell Research. 29 (1): 54–66. doi:10.1038/s41422-018-0104-9. PMC 6318295. PMID 30425322.
- ↑ Klemm SL, Shipony Z, Greenleaf WJ (April 2019). "Chromatin accessibility and the regulatory epigenome". Nature Reviews. Genetics. 20 (4): 207–220. doi:10.1038/s41576-018-0089-8. PMID 30675018. S2CID 59159906.
- ↑ Vongs A, Kakutani T, Martienssen RA, Richards EJ (June 1993). "Arabidopsis thaliana DNA methylation mutants". Science. 260 (5116): 1926–8. Bibcode:1993Sci...260.1926V. doi:10.1126/science.8316832. PMID 8316832.
- 1 2 Jeddeloh JA, Stokes TL, Richards EJ (May 1999). "Maintenance of genomic methylation requires a SWI2/SNF2-like protein". Nature Genetics. 22 (1): 94–7. doi:10.1038/8803. PMID 10319870. S2CID 20199014.
- ↑ Kankel MW, Ramsey DE, Stokes TL, Flowers SK, Haag JR, Jeddeloh JA, et al. (March 2003). "Arabidopsis MET1 cytosine methyltransferase mutants". Genetics. 163 (3): 1109–22. doi:10.1093/genetics/163.3.1109. PMC 1462485. PMID 12663548.
- ↑ Jones L, Ratcliff F, Baulcombe DC (May 2001). "RNA-directed transcriptional gene silencing in plants can be inherited independently of the RNA trigger and requires Met1 for maintenance". Current Biology. 11 (10): 747–57. doi:10.1016/s0960-9822(01)00226-3. PMID 11378384. S2CID 16789197.
- ↑ Chan SW, Henderson IR, Jacobsen SE (May 2005). "Gardening the genome: DNA methylation in Arabidopsis thaliana". Nature Reviews. Genetics. 6 (5): 351–60. doi:10.1038/nrg1601. PMID 15861207. S2CID 20083628.
- ↑ Li Y, Kumar S, Qian W (January 2018). "Active DNA demethylation: mechanism and role in plant development". Plant Cell Reports. 37 (1): 77–85. doi:10.1007/s00299-017-2215-z. PMC 5758694. PMID 29026973.
- ↑ Choi Y, Gehring M, Johnson L, Hannon M, Harada JJ, Goldberg RB, et al. (July 2002). "DEMETER, a DNA glycosylase domain protein, is required for endosperm gene imprinting and seed viability in arabidopsis". Cell. 110 (1): 33–42. doi:10.1016/s0092-8674(02)00807-3. PMID 12150995. S2CID 14828646.
- ↑ Zhu J, Kapoor A, Sridhar VV, Agius F, Zhu JK (January 2007). "The DNA glycosylase/lyase ROS1 functions in pruning DNA methylation patterns in Arabidopsis". Current Biology. 17 (1): 54–9. doi:10.1016/j.cub.2006.10.059. PMID 17208187. S2CID 3955783.
- ↑ Williams BP, Gehring M (December 2017). "Stable transgenerational epigenetic inheritance requires a DNA methylation-sensing circuit". Nature Communications. 8 (1): 2124. Bibcode:2017NatCo...8.2124W. doi:10.1038/s41467-017-02219-3. PMC 5730562. PMID 29242626.
- ↑ Wang J, Blevins T, Podicheti R, Haag JR, Tan EH, Wang F, Pikaard CS (August 2017). "Arabidopsis SMC4 identifies condensin as a corepressor of pericentromeric transposons and conditionally expressed genes". Genes & Development. 31 (15): 1601–1614. doi:10.1101/gad.301499.117. PMC 5630024. PMID 28882854.
- ↑ Córdoba-Cañero D, Cognat V, Ariza RR, Roldán Arjona T, Molinier J (December 2017). "Dual control of ROS1-mediated active DNA demethylation by DNA damage-binding protein 2 (DDB2)". The Plant Journal. 92 (6): 1170–1181. doi:10.1111/tpj.13753. PMID 29078035. S2CID 37919309.
- 1 2 3 4 Ream TS, Haag JR, Wierzbicki AT, Nicora CD, Norbeck AD, Zhu JK, et al. (January 2009). "Subunit compositions of the RNA-silencing enzymes Pol IV and Pol V reveal their origins as specialized forms of RNA polymerase II". Molecular Cell. 33 (2): 192–203. doi:10.1016/j.molcel.2008.12.015. PMC 2946823. PMID 19110459.
- 1 2 3 Huang Y, Kendall T, Forsythe ES, Dorantes-Acosta A, Li S, Caballero-Pérez J, et al. (July 2015). "Ancient Origin and Recent Innovations of RNA Polymerase IV and V". Molecular Biology and Evolution. 32 (7): 1788–99. doi:10.1093/molbev/msv060. PMC 4476159. PMID 25767205.
- ↑ Tucker SL, Reece J, Ream TS, Pikaard CS (2010). "Evolutionary history of plant multisubunit RNA polymerases IV and V: subunit origins via genome-wide and segmental gene duplications, retrotransposition, and lineage-specific subfunctionalization". Cold Spring Harbor Symposia on Quantitative Biology. 75: 285–97. doi:10.1101/sqb.2010.75.037. PMID 21447813.
- ↑ Luo J, Hall BD (January 2007). "A multistep process gave rise to RNA polymerase IV of land plants". Journal of Molecular Evolution. 64 (1): 101–12. Bibcode:2007JMolE..64..101L. doi:10.1007/s00239-006-0093-z. PMID 17160640. S2CID 37590716.
- 1 2 Haag JR, Brower-Toland B, Krieger EK, Sidorenko L, Nicora CD, Norbeck AD, et al. (October 2014). "Functional diversification of maize RNA polymerase IV and V subtypes via alternative catalytic subunits". Cell Reports. 9 (1): 378–390. doi:10.1016/j.celrep.2014.08.067. PMC 4196699. PMID 25284785.
- ↑ Ma L, Hatlen A, Kelly LJ, Becher H, Wang W, Kovarik A, et al. (September 2015). "Angiosperms Are Unique among Land Plant Lineages in the Occurrence of Key Genes in the RNA-Directed DNA Methylation (RdDM) Pathway". Genome Biology and Evolution. 7 (9): 2648–62. doi:10.1093/gbe/evv171. PMC 4607528. PMID 26338185.
- ↑ Yaari R, Katz A, Domb K, Harris KD, Zemach A, Ohad N (April 2019). "RdDM-independent de novo and heterochromatin DNA methylation by plant CMT and DNMT3 orthologs". Nature Communications. 10 (1): 1613. Bibcode:2019NatCo..10.1613Y. doi:10.1038/s41467-019-09496-0. PMC 6453930. PMID 30962443.
- 1 2 Moran Y, Agron M, Praher D, Technau U (February 2017). "The evolutionary origin of plant and animal microRNAs". Nature Ecology & Evolution. 1 (3): 27. Bibcode:2017NatEE...1...27M. doi:10.1038/s41559-016-0027. PMC 5435108. PMID 28529980.
- ↑ Castel SE, Martienssen RA (February 2013). "RNA interference in the nucleus: roles for small RNAs in transcription, epigenetics and beyond". Nature Reviews. Genetics. 14 (2): 100–12. doi:10.1038/nrg3355. PMC 4205957. PMID 23329111.
- ↑ Volpe TA, Kidner C, Hall IM, Teng G, Grewal SI, Martienssen RA (September 2002). "Regulation of heterochromatic silencing and histone H3 lysine-9 methylation by RNAi". Science. 297 (5588): 1833–7. Bibcode:2002Sci...297.1833V. doi:10.1126/science.1074973. PMID 12193640. S2CID 2613813.
- ↑ Bühler M, Verdel A, Moazed D (June 2006). "Tethering RITS to a nascent transcript initiates RNAi- and heterochromatin-dependent gene silencing". Cell. 125 (5): 873–86. doi:10.1016/j.cell.2006.04.025. PMID 16751098. S2CID 2938057.
- ↑ Zaratiegui M, Castel SE, Irvine DV, Kloc A, Ren J, Li F, et al. (October 2011). "RNAi promotes heterochromatic silencing through replication-coupled release of RNA Pol II". Nature. 479 (7371): 135–8. Bibcode:2011Natur.479..135Z. doi:10.1038/nature10501. PMC 3391703. PMID 22002604.
- ↑ Fagard M, Vaucheret H (June 2000). "(TRANS)GENE SILENCING IN PLANTS: How Many Mechanisms?". Annual Review of Plant Physiology and Plant Molecular Biology. 51: 167–194. doi:10.1146/annurev.arplant.51.1.167. PMID 15012190.
- ↑ Napoli C, Lemieux C, Jorgensen R (April 1990). "Introduction of a Chimeric Chalcone Synthase Gene into Petunia Results in Reversible Co-Suppression of Homologous Genes in trans". The Plant Cell. 2 (4): 279–289. doi:10.1105/tpc.2.4.279. PMC 159885. PMID 12354959.
- ↑ van der Krol AR, Mur LA, Beld M, Mol JN, Stuitje AR (April 1990). "Flavonoid genes in petunia: addition of a limited number of gene copies may lead to a suppression of gene expression". The Plant Cell. 2 (4): 291–9. doi:10.1105/tpc.2.4.291. PMC 159886. PMID 2152117.
- ↑ Depicker A, Montagu MV (June 1997). "Post-transcriptional gene silencing in plants". Current Opinion in Cell Biology. 9 (3): 373–82. doi:10.1016/s0955-0674(97)80010-5. PMID 9159078.
- ↑ Assaad FF, Tucker KL, Signer ER (September 1993). "Epigenetic repeat-induced gene silencing (RIGS) in Arabidopsis". Plant Molecular Biology. 22 (6): 1067–85. doi:10.1007/BF00028978. PMID 8400126. S2CID 26576784.
- ↑ Ingelbrecht I, Van Houdt H, Van Montagu M, Depicker A (October 1994). "Posttranscriptional silencing of reporter transgenes in tobacco correlates with DNA methylation". Proceedings of the National Academy of Sciences of the United States of America. 91 (22): 10502–6. Bibcode:1994PNAS...9110502I. doi:10.1073/pnas.91.22.10502. PMC 45049. PMID 7937983.
- ↑ Meyer P, Heidmann I (May 1994). "Epigenetic variants of a transgenic petunia line show hypermethylation in transgene DNA: an indication for specific recognition of foreign DNA in transgenic plants". Molecular & General Genetics. 243 (4): 390–9. doi:10.1007/BF00280469. PMID 8202084. S2CID 10429039.
- ↑ Greenberg MV, Ausin I, Chan SW, Cokus SJ, Cuperus JT, Feng S, et al. (March 2011). "Identification of genes required for de novo DNA methylation in Arabidopsis". Epigenetics. 6 (3): 344–54. doi:10.4161/epi.6.3.14242. PMC 3092683. PMID 21150311.
- ↑ Meyer P (2013). "Transgenes and their contributions to epigenetic research". The International Journal of Developmental Biology. 57 (6–8): 509–15. doi:10.1387/ijdb.120254pm. PMID 24166433.
- ↑ Hamilton AJ, Baulcombe DC (October 1999). "A species of small antisense RNA in posttranscriptional gene silencing in plants". Science. 286 (5441): 950–2. doi:10.1126/science.286.5441.950. PMID 10542148.
- 1 2 Mette MF, Aufsatz W, van der Winden J, Matzke MA, Matzke AJ (October 2000). "Transcriptional silencing and promoter methylation triggered by double-stranded RNA". The EMBO Journal. 19 (19): 5194–201. doi:10.1093/emboj/19.19.5194. PMC 302106. PMID 11013221.
- ↑ Hamilton A, Voinnet O, Chappell L, Baulcombe D (September 2002). "Two classes of short interfering RNA in RNA silencing". The EMBO Journal. 21 (17): 4671–9. doi:10.1093/emboj/cdf464. PMC 125409. PMID 12198169.
- 1 2 Xie Z, Johansen LK, Gustafson AM, Kasschau KD, Lellis AD, Zilberman D, et al. (May 2004). "Genetic and functional diversification of small RNA pathways in plants". PLOS Biology. 2 (5): E104. doi:10.1371/journal.pbio.0020104. PMC 350667. PMID 15024409.
- ↑ Zilberman D, Cao X, Jacobsen SE (January 2003). "ARGONAUTE4 control of locus-specific siRNA accumulation and DNA and histone methylation". Science. 299 (5607): 716–9. Bibcode:2003Sci...299..716Z. doi:10.1126/science.1079695. PMID 12522258. S2CID 8498615.
- ↑ Dalmay T, Hamilton A, Rudd S, Angell S, Baulcombe DC (May 2000). "An RNA-dependent RNA polymerase gene in Arabidopsis is required for posttranscriptional gene silencing mediated by a transgene but not by a virus". Cell. 101 (5): 543–53. doi:10.1016/s0092-8674(00)80864-8. PMID 10850496. S2CID 2103803.
- ↑ Herr AJ, Jensen MB, Dalmay T, Baulcombe DC (April 2005). "RNA polymerase IV directs silencing of endogenous DNA". Science. 308 (5718): 118–20. Bibcode:2005Sci...308..118H. doi:10.1126/science.1106910. PMID 15692015. S2CID 206507767.
- ↑ Onodera Y, Haag JR, Ream T, Costa Nunes P, Pontes O, Pikaard CS (March 2005). "Plant nuclear RNA polymerase IV mediates siRNA and DNA methylation-dependent heterochromatin formation". Cell. 120 (5): 613–22. doi:10.1016/j.cell.2005.02.007. PMID 15766525. S2CID 1695604.
- ↑ Kanno T, Huettel B, Mette MF, Aufsatz W, Jaligot E, Daxinger L, et al. (July 2005). "Atypical RNA polymerase subunits required for RNA-directed DNA methylation". Nature Genetics. 37 (7): 761–5. doi:10.1038/ng1580. PMID 15924141. S2CID 20032369.
- ↑ Pontier D, Yahubyan G, Vega D, Bulski A, Saez-Vasquez J, Hakimi MA, et al. (September 2005). "Reinforcement of silencing at transposons and highly repeated sequences requires the concerted action of two distinct RNA polymerases IV in Arabidopsis". Genes & Development. 19 (17): 2030–40. doi:10.1101/gad.348405. PMC 1199573. PMID 16140984.
- 1 2 3 Bond DM, Baulcombe DC (January 2015). "Epigenetic transitions leading to heritable, RNA-mediated de novo silencing in Arabidopsis thaliana". Proceedings of the National Academy of Sciences of the United States of America. 112 (3): 917–22. Bibcode:2015PNAS..112..917B. doi:10.1073/pnas.1413053112. PMC 4311854. PMID 25561534.
- ↑ Kanazawa A, Inaba JI, Shimura H, Otagaki S, Tsukahara S, Matsuzawa A, et al. (January 2011). "Virus-mediated efficient induction of epigenetic modifications of endogenous genes with phenotypic changes in plants". The Plant Journal. 65 (1): 156–168. doi:10.1111/j.1365-313X.2010.04401.x. hdl:2115/49399. PMID 21175898.
- ↑ Dalakouras A, Moser M, Zwiebel M, Krczal G, Hell R, Wassenegger M (December 2009). "A hairpin RNA construct residing in an intron efficiently triggered RNA-directed DNA methylation in tobacco". The Plant Journal. 60 (5): 840–51. doi:10.1111/j.1365-313X.2009.04003.x. PMID 19702668.
- ↑ Pignatta D, Novitzky K, Satyaki PR, Gehring M (November 2018). "A variably imprinted epiallele impacts seed development". PLOS Genetics. 14 (11): e1007469. doi:10.1371/journal.pgen.1007469. PMC 6237401. PMID 30395602.
- ↑ Papikian A, Liu W, Gallego-Bartolomé J, Jacobsen SE (February 2019). "Site-specific manipulation of Arabidopsis loci using CRISPR-Cas9 SunTag systems". Nature Communications. 10 (1): 729. Bibcode:2019NatCo..10..729P. doi:10.1038/s41467-019-08736-7. PMC 6374409. PMID 30760722.
- ↑ Dalakouras A, Wassenegger M, Dadami E, Ganopoulos I, Pappas ML, Papadopoulou K (January 2020). "Genetically Modified Organism-Free RNA Interference: Exogenous Application of RNA Molecules in Plants". Plant Physiology. 182 (1): 38–50. doi:10.1104/pp.19.00570. PMC 6945881. PMID 31285292.
- ↑ Regalado A (11 August 2015). "The Next Great GMO Debate". MIT Technology Review.
- ↑ Gohlke J, Mosher RA (September 2015). "Exploiting mobile RNA silencing for crop improvement". American Journal of Botany. 102 (9): 1399–400. doi:10.3732/ajb.1500173. PMID 26391704.