| Pestivirus | |
|---|---|
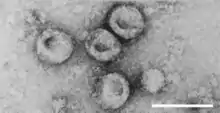 | |
| Virions of Pestivirus sp. | |
| Virus classification | |
| (unranked): | Virus |
| Realm: | Riboviria |
| Kingdom: | Orthornavirae |
| Phylum: | Kitrinoviricota |
| Class: | Flasuviricetes |
| Order: | Amarillovirales |
| Family: | Flaviviridae |
| Genus: | Pestivirus |
| Species | |
| |
Pestivirus is a genus of viruses, in the family Flaviviridae. Viruses in the genus Pestivirus infect mammals, including members of the family Bovidae (which includes cattle, sheep, and goats) and the family Suidae (which includes various species of swine). There are 11 species in this genus. Diseases associated with this genus include: hemorrhagic syndromes, abortion, and fatal mucosal disease.[1][2]
Structure
Viruses in Pestivirus are enveloped, with spherical geometries. Their diameter is around 50 nm. Genomes are linear and not segmented, around 12kb in length.[1]
| Genus | Structure | Symmetry | Capsid | Genomic arrangement | Genomic segmentation |
|---|---|---|---|---|---|
| Pestivirus | Icosahedral-like | Pseudo T=3 | Enveloped | Linear | Monopartite |
Lifecycle
Entry into the host cell is achieved by attachment of the viral envelope protein E2 to host receptors, which mediates clathrin-mediated endocytosis. The main viral replication process happens in host cytoplasm. Replication follows the positive strand RNA virus replication model. An IRES RNA element at the 5'-nontranslated region (NTR) of the viral genome recruits viral and cellular translation factors to initiate viral protein translation.[3] Viral proteins are first translated as polyprotein, and then processed into individual structure and non-structure proteins by both viral and host proteases.[3] The virus exits the host cell by budding. Mammals serve as the natural hosts.When infected, the host sheds viruses in almost all body secretions including saliva, nasal discharge, milk, and feaces.[3] Vertical transmission (viruses crossing the placenta and infecting the fetus) are also common.[1]
| Genus | Host details | Tissue tropism | Entry details | Release details | Replication site | Assembly site | Transmission |
|---|---|---|---|---|---|---|---|
| Pestivirus | Mammals | None | Clathrin-mediated endocytosis | Secretion | Cytoplasm | Cytoplasm | Horizontal and Vertical |
Genome
Pestivirus viruses have a single strand of positive-sense RNA (i.e. RNA which can be directly translated into viral proteins) that is around 12.5 kilobases (kb) long (equal to the length of 12,500 nucleotides), but due to recombination events has been observed up to 16.5 kilobases in length.[4] Sometimes, virions (individual virus particles) contain sections of an animal's genome that have been duplicated, though this is not normally the case. Although lacking Poly-A tail at the 3' end of the genome, it contains stem-loop regions that might be involved in viral translation and replication.[5] The genome contains RNA to encode both structural and nonstructural proteins. The molecular biology of pestiviruses shares many similarities and peculiarities with the human hepaciviruses. Genome organisation and translation strategy are highly similar for the members of both genera. For BVDV, frequently nonhomologous RNA recombination events lead to the appearance of genetically distinct viruses that are lethal to the host.[6]
Transmission and prevention
Pestivirus A is widespread in Australia, mainly in cattle. Some adult cattle are immune to the disease, while others are lifelong carriers. If a foetus becomes infected within the first three to four months of gestation, then it will fail to develop antibodies towards the virus. In these cases, the animals often die before birth or shortly after. It is spread very easily among feedlot cattle as nasal secretions and close contact spread the disease, and animals with infected mucous membranes give off millions of particles of BVDV a day.
Symptoms of Pestivirus infection include diarrhoea, respiratory problems, and bleeding disorders.
Pestivirus A vaccines exist and the correct vaccine strain should be given, depending on the herd's location and the endemic strain in that region. This vaccination must be given regularly to maintain immunity.
Vaccines
There are 120 registered BVD vaccine products currently used around the world, mainly in North and South America.[7] These are conventional modified live virus (MLV) or inactivated/killed virus vaccines.[7] In pregnant animals live vaccines pose significant risk of vertical transmission of vaccine virus that can occasionally result in complications for calves.[8] Most of the harm done by BVDV is to unborn calves and depends on the timing of infection.[9] Vaccination has not proved to be effective for Bovine Viral Diarrhea (BVD), as the presence of BVD has not lessened since the vaccine has been developed.[10] Animals who are affected by the virus during early fetal development may become persistently infected (PI) and lack an immune response to BVD. These animal’s presence in herds and them shedding virus can infect other animals in the herd before vaccination is possible.[11] PI animals do not produce antibodies and are the main source of infection for herds, so culling is necessary to eradicate infection sources.[3] Vaccines are not able to prevent fetal infections, so this poses a huge source of infection for cattle herds.[10] Another reason for the inefficiency of the BVD vaccine is because of failure to vaccinate whole areas, rather than just individual herds.[11] Border Disease, which affects lambs, is also caused by Pestivirus, but has no vaccine at this time.[12] Marker vaccines are beneficial tools for the eradication of animal diseases in regions with a high prevalence of the designated disease. The chimeric CP7_E2alf used to see how altered cell tropism affects pigs may not only serve as a tool for a better understanding of Pestivirus attachment, entry, and assembly, but also represent modified live CSFV "marker vaccines."[3]
Structural and non-structural proteins
Genomic RNA of pestiviruses is translated into a large polyprotein that is divided into several proteins. It has a single big open reading frame (ORF) that can encode roughly 4000 amino acids and a positive-sense ssRNA genome. Among the structural proteins that are N terminal in this polyprotein are three glycoproteins, which are referred to as E0, E1, and E2 depending on the order in which they end up appearing in the polyprotein.[13] The nucleocapsid protein C and the three envelope glycoproteins Erns, E1, and E2 are the virion's structural components.[14] Beginning with a nascent cleavage between the precursor ErnsE1E2 and the capsid protein, glycoprotein processing is then carried out by cleavage at the C-terminal end of E2.[14] After being split into ErnsE1 and E2, ErnsE1 is then transformed into Erns and E1. A host signal peptidase located in the endoplasmic reticulum's lumen catalyzes the cleavage between Erns and E1, as well as that between E1 and E2 (ER).[15] A new type of signal peptidase cleavage site is identified in an RNA virus polyprotein. The most important structural protein is E2, which regulates cell tropism by interacting with cell surface receptors and inducing responses from cytotoxic T-lymphocytes and neutralizing antibodies. E2 is a type I transmembrane protein and has a mass of 55 kDa. All three glycoproteins aid in the attachment of the virus and its entry into target cells. Viral entry and contagiousness require heterodimeric E1-E2 molecules. E1 is categorized as a type I transmembrane protein and has a mass of 33 kDa. Of the three glycoproteins, the functions of E1 are the least developed and least understood.[16] A virus's glycoproteins must perform a variety of tasks throughout its life cycle in order for the virus to successfully infect cells or animals, multiply, and then leave the affected cells. These activities can be broken down into the three mutually exclusive categories of interacting with hosts to sustain itself throughout the animal population, interacting with cells to infect and replicate, and connecting with other viral proteins to form viable virions. Although it lacks a hydrophobic anchor sequence, the structural glycoprotein E(rns) of pestiviruses has been found to be connected to the virion and to membranes in infected cells via its COOH terminus. Erns, an envelope glycoprotein, was recently recognized as an RNase. RNases have a variety of biological effects. They have been proven to be immunosuppressive, neurotoxic, and antihelminthic. Erns severely reduced the protein synthesis of various kinds of lymphocytes without causing cell membrane damage.[17] Symptoms of pestivirus infections include leukopenia and immunosuppression. In the pathogenesis of pestiviruses, ERNS is crucial. A pestivirus envelope glycoprotein called ERNS is crucial for virus attachment and cell infection. Erns lacks a transmembrane domain, unlike the other two envelope proteins E1 and E2, and a significant amount is secreted into the medium of infected. Erns's C-terminus serves as a membrane anchor, a retention/secretion signal, a binding site for cell surface glycosaminoglycans (GAGs), a signal peptidase cleavage site, and more. Erns has a mass of 44–48 kDa.[18] The protein is also present in some pure pestivirus virions, which begs the crucial and fascinating question of how it attaches to the pestivirus envelope. Virus-neutralizing antibodies primarily target the pestivirus E2 glycoproteins, which also function in receptor binding and host range limiting. At the moment where pestiviruses enter cells, their host specificity is probably influenced by the sequence and structure of E2. Enveloped viruses have created a variety of crafty invasion methods.[19] For cell attachment and membrane fusion to occur, one or more viral envelope glycoproteins are required. In contrast to pestiviruses and hepacivirus, which both have two envelope glycoproteins, E1 and E2, members of the Flaviviridae family, such as flaviviruses, only have one glycoprotein, E, in their envelope. Although E2 participates in cell attachment, it is not yet known which protein causes membrane fusion.[20]
The bovine viral diarrhea virus (BVDV) is what causes bovine viral diarrhea (BVD). Bovine viral diarrhea virus type 1 (BVDV-1), Bovine viral diarrhea virus type 2 (BVDV-2), Border disease virus (BDV), and Classical swine fever (CSF) virus are the four recognized species in the genus Pestivirus of the family Flaviviridae.[21] Although progress has been made in recent decades in identifying the activities of the BVDV NSPs, research on the virus still mostly focuses on its structural protein. Understanding BVDV non-structural proteins would assist researchers to better comprehend viral replication and the molecular basis of viral persistent infection. Eight non-structural proteins (NSPs) are encoded by the bovine viral diarrhea virus (BVDV) (i.e., Npro, p7, NS2, NS3, NS4A, NS4B, NS5A, and NS5B). A single open reading frame is encoded by a singular, single-stranded, positive-stranded RNA of 12.3–16.5 kb in the BVDV (ORF). The coding sequence is NH2, and the ORF can be split into various parts to encode polyproteins. –Npro (p20) (p20) –C (p14) (p14) -Erns/E0(gp48), -E1(gp25), -E2(gp53), -p7, NS2(p54), -NS3(p80), -NS4A(p10), -NS4B(p30), -NS5A(p58), -NS5B(p75), -COOH. Individually or collectively, these proteins are involved in viral replication, transcription, and translation. Npro (p20), a protein specific to pestivirus with a molecular weight of roughly 20 kDa, is the first protein generated from the N-terminus of the viral polyprotein. BVDV Npro is a hydrophilic outer membrane protein that primarily consists of beta-sheets and random curling.[22] It lacks a signal peptide. Npro is also a self-protease that can catalyze the breakdown of developing polyproteins to create the BVDV C protein. Infected animals have innate immune suppression as a result of BVDV Npro's capacity to control the generation or inhibition of type I interferon (IFN-I) and alter the virus' ability to replicate. A 6-7 kDa polypeptide generated from E2 called viral protein p7 has two domains. The other domain, which is present throughout infection in the cell as free p7 or E2-p7, is released by signal peptidase interpretation and is found at the C-terminus of E2 without being cleaved. However, because p7 was not found in BVDV particles, it was categorized as a non-structural protein. Although BVDV p7 can aid in the production of contagious BVDV particles and encourage virus release, the exact mechanisms behind these actions are still unknown.[23] With 450 amino acids, NS2 (p54) is a cysteine protease. A shared domain of the C-terminal protease structure and a hydrophobic N-terminal half-anchored protein membrane make up this structure.[24] NS2-NS3 cleavage is mediated by the self-protease in NS2, which may effectively cleave into NS2 and NS3 in the early stages of infection, and the degree of NS2-NS3 cleavage controls BVDV from RNA replication to morphological alterations.[25] Additionally, when the BVDV virus infects a cell, the cell chaperone DNAJC14 joins forces with the viral NS2-NS3 to facilitate the activation of the NS2 protease and the release of NS3, which facilitates the production of virions.[26] As a target antigen for ELISA BVDV detection, NS3 is a multifunctional protein with serine protease activity, helicase activity, and nucleoside triphosphatase (NTPase) activity.[13] Although it plays a significant role in the BVDV replicase and controls the viral RNAs ability to replicate, NS3 has little impact on the assembly of the virus. Only in the NS3/NS4A complex can the NS3 protease reach peak activity, after which the C-terminus of NS3 cleaves all downstream proteins. The replication of viral RNA will be hampered by the inactivation of the NS3 protease, helicase, and NTPase. Normal detection limits for the NS2-NS3 (p125) protein in Ncp and Cp BVDV-infected cells are 120 kDa. The cleavage of NS2-NS3 is connected to the replication of the virus in the early stages of virus infection.[13] A complex known as NS2-NS3/NS4A (NS2-3/4A) is created when NS4A joins with uncleaved NS2-NS3 (NS2-3) or NS3/NS4A. It can be utilized to support RNA replication and virus assembly as the fundamental element of virus particles. In the NS3/NS4A serine protease complex, NS4A functions as a protease cofactor, engaging with NS3 to catalyze the cleavage of downstream proteins NS4B, NS5A, and NS5B.[27] In particle assembly, NS2 and NS3 can replace uncut NS2-NS3 molecules, but the precise mechanism is still unknown.[13] A 35 kDa hydrophobic protein with NTPase activity called NS4B (p30) is involved in the replication of the BVDV genome.[28] Due to interactions between the viral Npro, Erns, and NS4B and the host immune signaling pathways, BVDV can bypass the host immune response and cause persistent infection in cattle by blocking their innate immune responses.[29] The primary target for the diagnosis of diseases, the creation of vaccines, and the management of infections is NS4B. After viral infection, NS4B can trigger humoral and cellular immune responses thanks to its highly conserved epitopes. NS5B (p75), which features a functional motif typical of viral RNA-dependent RNA polymerase, is roughly 77 kDa in size (RdRp). It primarily participates in the process of virus-infected cell membrane rearrangement and catalyzes the creation of viral RNA.[30] The C-terminus of the BVDV polyprotein is where the NS5A (p58) and NS5B (p75) are separated. Infected cells typically contain NS5A (p58) as a single protein or as an uncleaved NS5A-NS5B complex. A hydrophilic, phosphorylated protein with a molecular weight of 58 kDa called NS5A is a part of the viral replicase.[31] Although NS5B has a significant impact on RNA replication, its lack of specificity may have an impact on the design of viral replicase.[32] A number of issues, including the pathogenic mechanism, the regulation of virus replication, and the interaction between p7, NS4B, NS5A, and other NSP, remain unresolved.[33]
Species
- Pestivirus A or Bovine viral diarrhea virus 1 (BVDV-1), causes Bovine viral diarrhea and Mucosal disease
- Pestivirus B or Bovine viral diarrhea virus 2 or (BVDV-2), causes Bovine viral diarrhea and Mucosal disease
- Pestivirus C or Classical swine fever virus (CSFV), causes Classical swine fever
- Pestivirus D or Border disease virus (BDV), causes Border disease
- Pestivirus E or pronghorn pestivirus
- Pestivirus F or Bungowannah virus
- Pestivirus G or giraffe pestivirus
- Pestivirus H or Hobi-like pestivirus
- Pestivirus I or Aydin-like pestivirus
- Pestivirus J or rat pestivirus
- Pestivirus K or atypical porcine pestivirus
- (Dongyang pangolin virus, DYPV)[34]
See also
References
- 1 2 3 "Viral Zone". ExPASy. Retrieved 15 June 2015.
- ↑ "Virus Taxonomy: 2020 Release". International Committee on Taxonomy of Viruses (ICTV). March 2021. Retrieved 16 May 2021.
- 1 2 3 4 5 Tautz N, Tews BA, Meyers G (2015). "The Molecular Biology of Pestiviruses". Advances in Virus Research. Academic Press. 93: 47–160. doi:10.1016/bs.aivir.2015.03.002. ISBN 9780128021798. PMID 26111586.
- ↑ Meyers G, Tautz N, Stark R, Brownlie J, Dubovi EJ, Collett MS, Thiel HJ (November 1992). "Rearrangement of viral sequences in cytopathogenic pestiviruses". Virology. 191 (1): 368–386. doi:10.1016/0042-6822(92)90199-Y. PMC 7131167. PMID 1329326.
- ↑ Pankraz A, Thiel HJ, Becher P (July 2005). "Essential and nonessential elements in the 3' nontranslated region of Bovine viral diarrhea virus". Journal of Virology. 79 (14): 9119–9127. doi:10.1128/JVI.79.14.9119-9127.2005. PMC 1168729. PMID 15994806.
- ↑ Rümenapf T, Thiel HJ (2008). "Molecular Biology of Pestiviruses". In Mettenleiter TC, Sobrino F (eds.). Animal Viruses: Molecular Biology. Caister Academic Press. ISBN 978-1-904455-22-6.
- 1 2 Riitho V, Strong R, Larska M, Graham SP, Steinbach F (October 2020). "Bovine Pestivirus Heterogeneity and Its Potential Impact on Vaccination and Diagnosis". Viruses. 12 (10): 1134. doi:10.3390/v12101134. PMC 7601184. PMID 33036281.
- ↑ Harasawa R (January 1995). "Adventitious pestivirus RNA in live virus vaccines against bovine and swine diseases". Vaccine. 13 (1): 100–103. doi:10.1016/0264-410X(95)80018-9. PMID 7762264.
- ↑ Laven, Richard (30 September 2010). "Diagnosis of bovine viral diarrhoea virus (BVDV)-associated problems". Livestock. 13 (3): 37–41. doi:10.1111/j.2044-3870.2008.tb00163.x.
- 1 2 Moennig V, Becher P (June 2015). "Pestivirus control programs: how far have we come and where are we going?". Animal Health Research Reviews. 16 (1): 83–87. doi:10.1017/S1466252315000092. PMID 26050577. S2CID 21890278.
- 1 2 Hamers C, Dehan P, Couvreur B, Letellier C, Kerkhofs P, Pastoret PP (March 2001). "Diversity among bovine pestiviruses". Veterinary Journal. 161 (2): 112–122. doi:10.1053/tvjl.2000.0504. PMID 11243683.
- ↑ Nettleton (1990). "Pestivirus infections in ruminants other than cattle". Revue Scientifique et Technique. 9 (1).
- 1 2 3 4 Zhou, Huan-Xiang (5 November 2008). "The debut of PMC Biophysics". PMC Biophysics. 1 (1): 1. doi:10.1186/1757-5036-1-1. ISSN 1757-5036. PMC 2605105. PMID 19351423.
- 1 2 Wright, K (July 1990). "Post-translational processing of the glycoproteins of lymphocytic choriomeningitis virus". Virology. 177 (1): 175–183. doi:10.1016/0042-6822(90)90471-3. ISSN 0042-6822. PMC 7130728. PMID 2141203.
- ↑ Bintintan, Ioana; Meyers, Gregor (March 2010). "A New Type of Signal Peptidase Cleavage Site Identified in an RNA Virus Polyprotein". Journal of Biological Chemistry. 285 (12): 8572–8584. doi:10.1074/jbc.M109.083394. PMC 2838279. PMID 20093364.
- ↑ Asfor, A.S.; Wakeley, P.R.; Drew, T.W.; Paton, D.J. (August 2014). "Recombinant pestivirus E2 glycoproteins prevent viral attachment to permissive and non permissive cells with different efficiency". Virus Research. 189: 147–157. doi:10.1016/j.virusres.2014.05.016. PMID 24874197.
- ↑ Wang, Jimin; Li, Yue; Modis, Yorgo (April 2014). "Structural models of the membrane anchors of envelope glycoproteins E1 and E2 from pestiviruses". Virology. 454–455: 93–101. doi:10.1016/j.virol.2014.02.015. PMC 3986810. PMID 24725935.
- ↑ Yu, Xinyou; Li, Tong; Li, Tianzhi; Dong, Lin; Wang, Jinliang; Shen, Zhiqiang (2022). "Establishment of a Dual SYBR Green I Fluorescence PCR Assay for African Swine Fever Virus and Porcine Epidemic Diarrhea Virus". Proceedings of the 8th International Conference on Agricultural and Biological Sciences. Shenzhen, China: SCITEPRESS - Science and Technology Publications. pp. 5–10. doi:10.5220/0011594000003430. ISBN 978-989-758-607-1. S2CID 252836671.
- ↑ Tews, Birke Andrea; Meyers, Gregor (November 2007). "The Pestivirus Glycoprotein Erns Is Anchored in Plane in the Membrane via an Amphipathic Helix". Journal of Biological Chemistry. 282 (45): 32730–32741. doi:10.1074/jbc.M706803200. PMID 17848558.
- ↑ Rümenapf, T; Unger, G; Strauss, J H; Thiel, H J (June 1993). "Processing of the envelope glycoproteins of pestiviruses". Journal of Virology. 67 (6): 3288–3294. doi:10.1128/jvi.67.6.3288-3294.1993. ISSN 0022-538X. PMC 237670. PMID 8388499.
- ↑ Nakamura, Shigeyuki; Fukusho, Akio; Inoue, Yoshimitsu; Sasaki, Hideharu; Ogawa, Nobuo (December 1993). "Isolation of different non-cytopathogenic bovine viral diarrhoea (BVD) viruses from cytopathogenic BVD virus stocks using reverse plaque formation method". Veterinary Microbiology. 38 (1–2): 173–179. doi:10.1016/0378-1135(93)90084-K. PMID 8128599.
- ↑ Wang, Shaokui; Li, Shan; Liu, Qian; Wu, Kun; Zhang, Jianqing; Wang, Shuansuo; Wang, Yi; Chen, Xiangbin; Zhang, Yi; Gao, Caixia; Wang, Feng; Huang, Haixiang; Fu, Xiangdong (August 2015). "The OsSPL16-GW7 regulatory module determines grain shape and simultaneously improves rice yield and grain quality". Nature Genetics. 47 (8): 949–954. doi:10.1038/ng.3352. ISSN 1061-4036. PMID 26147620. S2CID 28088659.
- ↑ Oestringer, Benjamin P.; Bolivar, Juan H.; Claridge, Jolyon K.; Almanea, Latifah; Chipot, Chris; Dehez, François; Holzmann, Nicole; Schnell, Jason R.; Zitzmann, Nicole (December 2019). "Hepatitis C virus sequence divergence preserves p7 viroporin structural and dynamic features". Scientific Reports. 9 (1): 8383. Bibcode:2019NatSR...9.8383O. doi:10.1038/s41598-019-44413-x. ISSN 2045-2322. PMC 6557816. PMID 31182749.
- ↑ Walther, Thomas; Bruhn, Barbara; Isken, Olaf; Tautz, Norbert (22 October 2021). "A novel NS3/4A protease dependent cleavage site within pestiviral NS2". Journal of General Virology. 102 (10). doi:10.1099/jgv.0.001666. ISSN 0022-1317. PMID 34676824. S2CID 239457986.
- ↑ Lattwein, E.; Klemens, O.; Schwindt, S.; Becher, P.; Tautz, N. (January 2012). "Pestivirus Virion Morphogenesis in the Absence of Uncleaved Nonstructural Protein 2-3". Journal of Virology. 86 (1): 427–437. doi:10.1128/JVI.06133-11. ISSN 0022-538X. PMC 3255886. PMID 22031952.
- ↑ Wu, Ming-Jhan; Shanmugam, Saravanabalaji; Welsch, Christoph; Yi, MinKyung (12 December 2019). James Ou, J.-H. (ed.). "Palmitoylation of Hepatitis C Virus NS2 Regulates Its Subcellular Localization and NS2-NS3 Autocleavage". Journal of Virology. 94 (1): e00906–19. doi:10.1128/JVI.00906-19. ISSN 0022-538X. PMC 6912101. PMID 31597774.
- ↑ Tautz, Norbert; Tews, Birke Andrea; Meyers, Gregor (2015), The Molecular Biology of Pestiviruses, Advances in Virus Research, vol. 93, Elsevier, pp. 47–160, doi:10.1016/bs.aivir.2015.03.002, ISBN 9780128021798, PMID 26111586, retrieved 3 December 2022
- ↑ Li, Guangyu; Adam, Awadalkareem; Luo, Huanle; Shan, Chao; Cao, Zengguo; Fontes-Garfias, Camila R.; Sarathy, Vanessa V.; Teleki, Cody; Winkelmann, Evandro R.; Liang, Yuejin; Sun, Jiaren; Bourne, Nigel; Barrett, Alan D. T.; Shi, Pei-Yong; Wang, Tian (28 November 2019). "An attenuated Zika virus NS4B protein mutant is a potent inducer of antiviral immune responses". npj Vaccines. 4 (1): 48. doi:10.1038/s41541-019-0143-3. ISSN 2059-0105. PMC 6883050. PMID 31815005.
- ↑ Shan, Yue; Tong, Zhao; Jinzhu, Ma; Yu, Liu; Zecai, Zhang; Chenhua, Wu; Wenjing, Huang; Siyu, Liu; Nannan, Chen; Siyu, Su; Tongtong, Bai; Jiang, Huang; Biaohui, Bai; Xin, Jin; Yulong, Zhou (September 2021). "Bovine viral diarrhea virus NS4B protein interacts with 2CARD of MDA5 domain and negatively regulates the RLR-mediated IFN-β production". Virus Research. 302: 198471. doi:10.1016/j.virusres.2021.198471. ISSN 0168-1702. PMID 34097933.
- ↑ Gladue, Douglas P.; Gavrilov, Boris K.; Holinka, Lauren G.; Fernandez-Sainz, Ignacio J.; Vepkhvadze, N.G.; Rogers, Kara; O'Donnell, Vivian; Risatti, Guillermo R.; Borca, Manuel V. (March 2011). "Identification of an NTPase motif in classical swine fever virus NS4B protein". Virology. 411 (1): 41–49. doi:10.1016/j.virol.2010.12.028. ISSN 0042-6822. PMID 21236462.
- ↑ Weiskircher, Erica; Aligo, Jason; Ning, Gang; Konan, Kouacou V (3 November 2009). "Bovine viral diarrhea virus NS4B protein is an integral membrane protein associated with Golgi markers and rearranged host membranes". Virology Journal. 6 (1): 185. doi:10.1186/1743-422x-6-185. ISSN 1743-422X. PMC 2777160. PMID 19887001. S2CID 16389031.
- ↑ Wu, Jiqin; Lu, Guoliang; Zhang, Bo; Gong, Peng (January 2015). "Perturbation in the Conserved Methyltransferase-Polymerase Interface of Flavivirus NS5 Differentially Affects Polymerase Initiation and Elongation". Journal of Virology. 89 (1): 249–261. doi:10.1128/jvi.02085-14. ISSN 0022-538X. PMC 4301151. PMID 25320292.
- ↑ Kuo Yang, LM; Tseng, PY; Liaw, CC; Zhang, LJ; Tsai, KC; Lin, ZH; Ho, HO; Kuo, YH (25 November 2015). "Sesquiterpenoids from Taiwanese Vernonia cinerea". Planta Medica. 81 (16). doi:10.1055/s-0035-1565538. ISSN 0032-0943.
- ↑ Loeffelholz MJ, Fenwick BW (January 2021). "Taxonomic Changes for Human and Animal Viruses, 2018 to 2020". Journal of Clinical Microbiology. 59 (2): e01932-20. doi:10.1128/JCM.01932-20. PMC 8111125. PMID 32848040.
External links
- The 7th ESVV Pestivirus Symposium Archived 14 May 2008 at the Wayback Machine—Uppsala, Sweden 2008
- Animal viruses
- Virus Pathogen Database and Analysis Resource (ViPR): Flaviviridae
- Viralzone: Pestivirus
- ICTV