| Ignacius | |
|---|---|
 | |
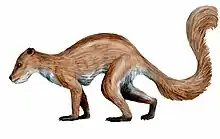
| Restoration of Ignacius graybullianus | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Order: | Plesiadapiformes |
| Family: | †Paromomyidae |
| Tribe: | †Phenacolemurini |
| Genus: | †Ignacius Matthew and Granger, 1921[1][2] |
| Type species | |
| †Ignacius frugivorus Matthew and Granger, 1921 | |
| Species | |
| |
Ignacius is a genus of extinct mammal from the early Cenozoic era. This genus is present in the fossil record from around 62-33 Ma (late Torrejonian-Chadronian North American Land Mammals Ages).[3][4] The earliest known specimens of Ignacius come from the Torrejonian of the Fort Union Formation, Wyoming[5] and the most recent known specimens from Ellesmere Island in northern Canada.[1] Ignacius is one of ten genera within the family Paromomyidae, the longest living family of any plesiadapiforms, persisting for around 30 Ma during the Paleocene and Eocene epochs.[6] The analyses of postcranial fossils by paleontologists suggest that members of the family Paromomyidae, including the genus Ignacius, most likely possessed adaptations for arboreality.[7]
Taxonomy
As of 2022, there were four valid species within the genus Ignacius: I. frugivorus,[8] I. fremontensis,[5] I. clarkforkensis,[7] and I. graybullianus.[3] There are also two described species of Ignacius from the Arctic of Canada.[1] The type species for the genus Ignacius is I. frugivorus and was found at the Mason Pocket locality in Colorado.[8] The holotype specimen (AMNH 17368), published in 1921 by Matthew and Granger, consists of an upper jaw with the canine, fourth premolar, first molar, and second molar.[8]
The genus Ignacius is situated within the family Paromomyidae and the order Plesiadapiformes. The relationship between plesiadapiforms and the modern group Euarchonta (Dermoptera, Scandentia, and Primates) is still under debate by paleontologists with some suggesting a closer relationship with primates,[7][9] and others suggesting a closer relationship with dermopterans.[10][11]
The genus Ignacius was found to be synonymous with the closely related genus, Phenacolemur, by G.G. Simpson in 1935[12] but revived by Bown and Rose in 1976.[3]
Description
Ignacius, along with the other plesiadapiforms, possess large, procumbent lower incisors convergent with those found in rodents, however, unlike rodents, the incisors of plesiadapiforms were not continuously growing.[3] Most species of Ignacius have a 1.0.1.3 lower dental formula although some specimens retain a small P3.[3] Compared with other paromomyids, Ignacius has small P4s in relation to M1s with relatively low crowned upper and lower molars.[3] The lower molars have reduced cusps, forming an almost continuous crest along the talonid basin.[9] The jaw of Ignacius is deep in comparison to tooth crown height, a feature distinct to this genus. Upper molars possess strong crests in a distinct ‘v’ shape between the paracone and metacone.[3]
Species
Ignacius frugivorus was the first species of Ignacius to be described, in 1921.[8] Ignacius frugivorus and I. fremontensis are the oldest species of Ignacius, both found in the Shotgun Local fauna of the Fort Union formation, Wyoming.[5] Until 2023,[1] the youngest known specimen of Ignacius was found in the Chadron formation of North Dakota and was represented by a single P4 (2). There is not enough material to identify this specimen to the species level.[4] Ignacius frugivorus and I. fremontensis are distinguishable from one another based primarily on the size of lower fourth premolars.[5] Ignacius fremontensis displays a significantly smaller P4 in relation to M1 when compared to I. frugivorus.[5] Ignacius clarkforkensis differs from other species in the retention of a single rooted P2 (5). Ignacius graybullianus has more quadrangular upper molars whereas the other three species are more triangular.[3] Strong v-shaped postparacone/prematacone cristae are present in I. graybullianus.[3] These cristae are less obliquely oriented in the other three taxa.[7] Ignacius mckennai and Ignacius dawsonae are the largest known species in the genus, living much further north and much later than the other known species, being found in Early Eocene deposits from Ellesmere Island.[1]
Paleoenvironment and geographic range
Ignacius occupied a broad geographic range during the early Cenozoic era in North America, ranging from Texas,[6][13] to the Canadian Arctic.[6][14] During the early Cenozoic, North America experienced multiple climatic changes with the warmest mean annual temperatures (around 16oC, 60oF) occurring during the Paleocene-Eocene Thermal Maximum (PETM) at around 55 Ma.[15] During the Eocene Climatic Optimum (c. 53 Ma) there was a correlated diversification of flora creating a complex, warm-temperate to subtropical habitat over much of interior North America.[16] The environment of Arctic Canada would have represented a temperate environment with winter temperatures rarely dropping below freezing.[14] The late Cretaceous and early Tertiary saw a diversification of angiosperms providing the opportunity for arboreal mammals to utilize the fruit, seed, exudate, and flower resources of these plants.[7] The environment of interior North America, extending into the Canadian Arctic, would have provided habitable ecosystems for these arboreal mammals to thrive and diversify.[7][14][16]
Functional morphology
Analysis of postcranial fossils of Ignacius suggest they were specialized for vertical climbing and clinging on tree trunks as well as for more agile arboreality than seen in other plesiadapiforms.[7][17] Some researchers have also hypothesized based on analysis of hand bone morphology, that they possessed gliding adaptations like those of modern-day flying lemurs (Order Dermoptera).[10] Further anatomical analysis of cranial synapomorphies, also suggest a close relationship between dermopterans and plesiadapiforms.[11]
Evidence for vertical climbing can be seen in the morphology of the humerus, femur, and tarsal bones which are consistent with increased flexibility of the shoulder, elbow, hip, knee, and ankle joints.[7][17] These features also suggest powerful flexion of the digits for grasping large tree trunks as well as small diameter support branches.[7][17] The features listed above are seen in many plesiadapiforms, but some anatomical features set paromomyids apart in being more adept for arboreal living. Flexible lumbar vertebrae as well as increased surface area on the innominate and femur for the origin and insertion of gluteal muscles, suggest paromomyids were capable of powerful bounding across tree branches.[7][17]
The large, procumbent incisors and reduced shearing crests of Paromomyids, especially Ignacius and Phenacolemur, suggest a diet specialized for feeding on exudates, comparable to the adaptions seen in extant callitrichine primates and Petaurus, a marsupial sugar-glider.[6][7][10][18]
References
- 1 2 3 4 5 Miller, K.; Tietjen, K.; Beard, K.C. (January 2023). "Basal Primatomorpha colonized Ellesmere Island (Arctic Canada) during the hyperthermal conditions of the early Eocene climatic optimum". PLOS ONE. 18 (1): e0280114. doi:10.1371/journal.pone.0280114. PMC 9876366. PMID 36696373.
- ↑ Matthew, W.D.; Granger, W. (September 1921). "New genera of Paleocene mammals" (PDF). American Museum Novitates (13): 1–7.
- 1 2 3 4 5 6 7 8 9 Bown, T. M., & Rose, K. D. (1976). New early Tertiary primates and a reappraisal of some Plesiadapiformes. Folia Primatologica, 26, 109– 138.
- 1 2 Kihm, A. J., & Tornow, M. A. (2014). First occurrence of plesiadapiform primates from the Chadronian (latest Eocene). Paludicola, 9, 176– 182.
- 1 2 3 4 5 Gazin, C. L. "Paleocene primates from the Shotgun Member of the Fort Union Formation in the Wind River Basin, Wyoming." Proceedings of the Biological Society of Washington 84, no. 3 (1971): 13-38.
- 1 2 3 4 Silcox, Mary T., Jonathan I. Bloch, Doug M. Boyer, Stephen GB Chester, and Sergi López‐Torres. "The evolutionary radiation of plesiadapiforms." Evolutionary Anthropology: Issues, News, and Reviews 26, no. 2 (2017): 74-94.
- 1 2 3 4 5 6 7 8 9 10 11 Bloch, J. I., Silcox, M. T., Boyer, D. M., & Sargis, E.J. (2007). New Paleocene skeletons and the relationship of “plesiadapiforms” to crownclade primates. Proceedings of the National Academy of Sciences, USA, 104, 1159–1164.
- 1 2 3 4 Matthew, William Diller, and Walter Granger. New genera of Paleocene mammals. By order of the Trustees of The American Museum of Natural History, 1921.
- 1 2 Silcox, M. T. (2008). The biogeographic origins of Primates and Euprimates: east, west, north, or south of Eden? In M. J. Dagosto, & E. J. Sargis (Eds.), Mammalian Evolutionary Morphology: A Tribute to Frederick S. Szalay (pp. 199–231). New York, NY: Springer-Verlag.
- 1 2 3 Beard, K. C. (1990). Gliding behavior and palaecology of the alleged primate family Paromomyidae (Mammalia, Dermoptera). Nature, 345, 340–341.
- 1 2 Kay, R. F., Thorington, R. W. Jr, Houde, P. (1990). Eocene plesiadapiform shows affinities with flying lemurs not primates. Nature, 345, 342– 344.
- ↑ Simpson, George Gaylord. "The Tiffany fauna, Upper Paleocene. 3, Primates, Carnivora, Condylarthra, and Amblypoda. American Museum novitates; no. 817." (1935).
- ↑ Schiebout, Judith Ann. Vertebrate paleontology and paleoecology of Paleocene Black Peaks Formation, Big Bend National Park, Texas. Texas Memorial Museum, The University of Texas at Austin, 1974.
- 1 2 3 Eberle, J. J., & Greenwood, D. R. (2012). Life at the top of the greenhouse Eocene world – A review of the Eocene flora and vertebrate fauna from Canada’s High Arctic. Geological Society of America Bulletin, 124,3.
- ↑ Wing, Scott L., Thomas M. Bown, and John D. Obradovich. "Early Eocene biotic and climatic change in interior western North America." Geology 19, no. 12 (1991): 1189-1192.
- 1 2 Woodburne, Michael O., Gregg F. Gunnell, and Richard K. Stucky. "Climate directly influences Eocene mammal faunal dynamics in North America." Proceedings of the National Academy of Sciences 106, no. 32 (2009): 13399-13403.
- 1 2 3 4 Bloch, J. I., & Boyer, D. M. (2007). New skeletons of Paleocene-Eocene Plesiadapiformes: a diversity of arboreal positional behaviors in early primates. In M. J. Ravosa, & M. Dagosto (Eds.), Primate Origins: Adaptations and Evolution (pp. 535–581). New York, NY: Springer.
- ↑ Kay, Richard F., and Matt Cartmill. "Cranial morphology and adaptations of Palaechthon nacimienti and other Paromomyidae (Plesiadapoidea,? Primates), with a description of a new genus and species." Journal of Human Evolution 6, no. 1 (1977): 19-53.
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||


.jpg.webp)