| Heat shock protein 104 | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Organism | |||||||
| Symbol | HSP104 | ||||||
| Entrez | 850633 | ||||||
| PDB | 5VJH | ||||||
| RefSeq (mRNA) | 1NM_001181846.1 | ||||||
| RefSeq (Prot) | NP_013074. | ||||||
| UniProt | P31539 | ||||||
| Other data | |||||||
| Chromosome | XII: 0.09 - 0.09 Mb | ||||||
| |||||||
Hsp104 is a heat-shock protein. It is known to reverse toxicity of mutant α-synuclein, TDP-43, FUS, and TAF15 in yeast cells.[1] Conserved in prokaryotes (ClpB), fungi, plants and aswell as animal mitochondria, there is yet to see hsp104 in multicellular animals. Hsp104 is classified as a. AAA+ ATPases and a subgroup of Hsp100/Clp, because of the usage of Atp hydrolysis for structural modulation of other proteins.[2] Hsp104 is not needed for normal cell growth but when exposed to stress there is an increase amount. Removing the aggregates without the hsp104 is insufficient there highlighting the importance of this heat shock protein and its interactions.[3]
Structure
Hsp104 monomer is composed of two NBDs (Nucleotide Binding Sites) NBD1 and NBD2 which communicate through allosteric communication . Located on the C-terminus of NBD1 there are around 125 residues that link both NBDs .In hsp104 NBD1 is where ATP hydrolysis occurs, NBD2 C-terminus is shown to express the configuration of the structure by nucleotide-dependent hexamerazation. These NBDs have diaphragms, which are conserved loops that interact in the middle of the channel inside the chaperone by coupling the ATP hydrolysis and polypeptide transport. Coupling of this reactions is needed due to without them coupled its an energetically unfavorable reaction.[4] These conserved loops contain conserved Tyr residues that are crucial for binding to substrates.[5]
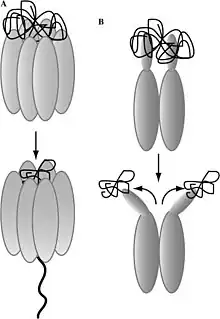
The Hsp104 homohexamer (Figure A) is a part of a unfolding/threading mechanism, that aggregates pass through and single polypeptides are extracted. Presence of ATP, ADP allows the formation of the Hsp104 monomers to homohexamer complexes. Through the hexamer complex the monomers communicate and make ATPhydrolysis from hsp104 hexarmerization. This ATP hydrolysis allows hsp104 to interact with substrates.[2]

Expression inducers
Stressors that induce Hsp104 expression include:[2]
- Heat or cold shock
- Hydrogen peroxide
- Ethanol
- Sodium Arsenide
Interactions
Prion propagation
The role of Hsp104 when interaction with Sup35 Prion Propagation is to interact with polymers. When Hsp104 is elevated it is hypothesized that amyloid polymers can be solubilized in misfolded monomers or intermediates, then later can be refolded into non prion monomers. When there is normal amount of Hsp104 the polymer breaks down into oligomeric “seeds”. Low levels of Hsp104 the polymer is not regulates forming aggregates within the cells that can lead to the cells death.[2]
Amyloid fibrils
No fibril formation is seen at a concentration of 1uM of Hsp104 At low concentrations of Hsp104 there is an observed concentration-dependent delay. A lag phase is seen when the Hsp104 concentration is at .01u. This control lag phase being 9 hours but after the low concentration of Hsp104 it is seen to be 72 hours. When the amount of Hsp104 increased the lag phase also increased inhibition of the Fibril formation.[6]
Dependencies of Hsp104
Hsp104 influences prion inheritance in yeast by remolding amyloids. When this happens prions infected show a cross-B structure and a amyloid fold.[7] Hsp104 has a chaperone called Hsp70 in yeast and is dependent for the full efficiency of thermotolarence. When Hsp70 is not accompanied by hsp104 it more useful than if both Hsp’s were gone. Hsp70 by itself can only do so much to regulate thermotolerance, hsp104 is neeeded when internal levels of Hsp70 are reducing. Hsp104 and Hsp70 also interact to recover native state of proteins that were exposed to heat and formed aggregates. When there is too much Hsp70/40 disaggragation activity by hsp104 increases. Small heat shock proteins (sHsps) are necessary for Hsp104 by helping with the clearance of aggregates. Some sHsps increase they’re molecular weight with partially folded proteins and are able to preform disaggragation by Hsp100/ClpB.[4]
References
- ↑ "Yeast Chaperone Melts Protein Aggregates". Alzforum.org. Retrieved September 12, 2016.
- 1 2 3 4 5 Romanova NV, Chernoff YO (2009). "Hsp104 and prion propagation". Protein and Peptide Letters. 16 (6): 598–605. doi:10.2174/092986609788490078. PMC 2791106. PMID 19519517.
- 1 2 Bösl B, Grimminger V, Walter S (October 2006). "The molecular chaperone Hsp104--a molecular machine for protein disaggregation". Journal of Structural Biology. 156 (1): 139–148. doi:10.1016/j.jsb.2006.02.004. PMID 16563798.
- 1 2 Grimminger-Marquardt V, Lashuel HA (March 2010). "Structure and function of the molecular chaperone Hsp104 from yeast". Biopolymers. 93 (3): 252–276. doi:10.1002/bip.21301. PMID 19768774.
- ↑ Shorter J, Southworth DR (August 2019). "Spiraling in Control: Structures and Mechanisms of the Hsp104 Disaggregase". Cold Spring Harbor Perspectives in Biology. 11 (8). doi:10.1101/cshperspect.a034033. PMC 6671941. PMID 30745294.
- ↑ Arimon M, Grimminger V, Sanz F, Lashuel HA (December 2008). "Hsp104 targets multiple intermediates on the amyloid pathway and suppresses the seeding capacity of Abeta fibrils and protofibrils". Journal of Molecular Biology. 384 (5): 1157–1173. doi:10.1016/j.jmb.2008.09.063. PMID 18851977.
- ↑ Desantis ME, Shorter J (January 2012). "The elusive middle domain of Hsp104 and ClpB: location and function". Biochimica et Biophysica Acta. 1823 (1): 29–39. doi:10.1016/j.bbamcr.2011.07.014. PMC 3219823. PMID 21843558.