
.PNG.webp)
The genetic history of Egypt reflects its geographical location at the crossroads of several major biocultural areas: North Africa, the Sahara, the Middle East, the Mediterranean and sub-Saharan Africa.
Genetic studies on ancient Egyptians
Egyptologist Barry Kemp has noted that DNA studies can only provide firm conclusions about the population of ancient Egypt if the sample results are of a significant number of individuals and represent a broad geographical and chronological range.[6] According to historian William Stiebling and archaeologist Susan N. Helft, conflicting DNA analysis on recent genetic samples such as the Amarna royal mummies has led to a lack of consensus on the genetic makeup of the ancient Egyptians and their geographic origins.[7]
2012 study of Ramesses III
In 2012, two mummies of two 20th dynasty individuals, Ramesses III and "Unknown Man E" believed to be Ramesses III's son Pentawer, were analyzed by Albert Zink, Yehia Z Gad, and a team of researchers under Zahi Hawass. Genetic kinship analyses revealed identical haplotypes in both mummies; using the Whit Athey's haplogroup predictor, the Y chromosomal haplogroup E1b1a was predicted.[8]
2017 DNA study of Mummies at Abusir el-Meleq
A study published in 2017 by Schuenemann et al. extracted DNA from 151 Egyptian mummies, whose remains were recovered from Abusir el-Meleq in Middle Egypt. The samples are from the time periods: Late New Kingdom, Ptolemaic, and Roman. Complete mtDNA sequences from 90 samples as well as genome-wide data from three ancient Egyptian individuals were successfully obtained and were compared with other ancient and modern datasets. The study used 135 modern Egyptian samples. The ancient Egyptian individuals in their own dataset possessed highly similar mtDNA haplogroup profiles, and cluster together, supporting genetic continuity across the 1,300-year transect. Modern Egyptians shared this mtDNA haplogroup profile, but also carried 8% more African component. A wide range of mtDNA haplogroups were found including clades of J, U, H, HV, M, R0, R2, K, T, L, I, N, X and W. In addition three ancient Egyptian individuals were analysed for Y-DNA, two were assigned to Middle Eastern haplogroup J and one to haplogroup E1b1b1a1b2. Both of these haplogroups are carried by modern Egyptians, and also common among Afroasiatic speakers in Northern Africa, Eastern Africa and the Middle East. The researchers cautioned that the examined ancient Egyptian specimens may not be representative of those of all ancient Egyptians since they were from a single archaeological site from the northern part of Egypt.[9] The analyses revealed that Ancient Egyptians had higher affinities with Near Eastern and European populations than do modern Egyptians, likely due to the 8% increase in the African component found in modern Egyptians.[9] However, comparative data from a contemporary population under Roman rule in Anatolia, did not reveal a closer relationship to the ancient Egyptians from the Roman period.[9] "Genetic continuity between ancient and modern Egyptians cannot be ruled out despite this more recent sub-Saharan African influx, while continuity with modern Ethiopians is not supported".[9]
The absolute estimates of sub-Saharan African ancestry in these three ancient Egyptian individuals ranged from 6 to 15%, and the absolute estimates of sub-Saharan African ancestry in the 135 modern Egyptian samples ranged from 14 to 21%, which show an 8% increase in African component. The age of the ancient Egyptian samples suggests that this 8% increase in African component occurred predominantly within the last 2000 years.[9] The 135 modern Egyptian samples were: 100 from modern Egyptians taken from a study by Pagani et al., and 35 from el-Hayez Western Desert Oasis taken from a study by Kujanova et al.[9] The 35 samples from el-Hayez Western Desert Oasis, whose population is described by the Kujanova et al. study as a mixed, relatively isolated, demographically small but autochthonous population, were already known from that study to have a relatively high sub-Saharan African component,[10] which is more than 11% higher than the African component in the 100 modern Egyptian samples.[11]
Verena Schuenemann and the authors of this study suggest a high level of genetic interaction with the Near East since ancient times, probably going back to Prehistoric Egypt although the oldest mummies at the site were from the New Kingdom: "Our data seem to indicate close admixture and affinity at a much earlier date, which is unsurprising given the long and complex connections between Egypt and the Middle East. These connections date back to Prehistory and occurred at a variety of scales, including overland and maritime commerce, diplomacy, immigration, invasion and deportation"[9]
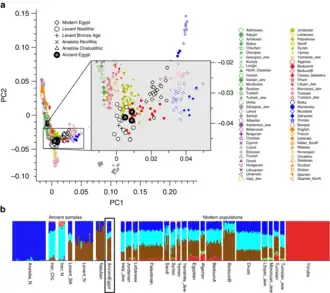 PCA and ADMIXTURE analysis of three ancient Egyptian samples and other modern and ancient populations.[9]
PCA and ADMIXTURE analysis of three ancient Egyptian samples and other modern and ancient populations.[9] PCA using only European samples based on the nuclear genome-wide data obtained on three ancient Egyptian samples.[9]
PCA using only European samples based on the nuclear genome-wide data obtained on three ancient Egyptian samples.[9] Complete results from the ADMIXTURE analysis using all samples in the merged data set, from the 2017 study by Schuenemann et al.[9]
Complete results from the ADMIXTURE analysis using all samples in the merged data set, from the 2017 study by Schuenemann et al.[9] FST values showing the genetic distances between 90 ancient Egyptians and modern populations. Blue values depict higher genetic distances, red values depict lower genetic distances between the ancient Egyptian population and modern populations in the respective area.[9]
FST values showing the genetic distances between 90 ancient Egyptians and modern populations. Blue values depict higher genetic distances, red values depict lower genetic distances between the ancient Egyptian population and modern populations in the respective area.[9]
Responses to the 2017 DNA study
The 2017 study has generated academic responses from scholars from other related disciplines, remarking on the conclusions of the study from a multi-disciplinary approach.
In 2021, Gourdine et al disputed Scheunemann et al's claim, in an unpublished article, that the increase in the sub-Saharan component in the modern Egyptian samples resulted from the trans-Saharan slave trade. Instead they argued that the sub-Saharan "genetic affinities" may be attributed to "early settlers" and "the relevant sub-Saharan genetic markers" do not correspond with the geography of known trade routes".[12]
In 2022, biological anthropologist S.O.Y. Keita argued that there were problems with the study's approaches and conclusions such as over-generalizations and a failure to consider alternative explanations. Particularly, he raised issues with the comparative samples from West Africa as a proxy group and generalisations about geographical Egypt and population origins from the sample results. He also drew attention to the fact that the authors draw inference on migrations in line with their Bayesian statistical approach rather than integrate other data into their explanations about the population history.[13]
In 2022, archaeologist Danielle Candelora stated that there were several limitations with the 2017 Scheunemann et al. study such as “new (untested) sampling methods, small sample size and problematic comparative data”.[14]
In 2023, Stiebling and Helft acknowledged that the 2017 study had performed the largest study on ancient Egyptians but noted that the findings still derived from a small sample of mummies from one site in Middle Egypt dating to the New Kingdom and later periods. They also stated that this study could not represent earlier populations or Egyptians from Upper Egypt who were geographically closer to Sub-Saharan populations.[15]
In 2023, Christopher Ehret argued that the conclusions of the 2017 study were based on insufficiently small sample sizes, and that the authors had a biased interpretation of the genetic data.[16] Ehret also criticised the Schuenemann article for asserting that there was “no sub-Saharan genetic component” in the Egyptian population and cited previous genetic analysis which had already identified the Horn of Africa as a source of a significant population component that spread into Egypt.[17]
Later findings
A unpublished, follow-up study by Schuenemann & Urban et al. (2021) was carried out collecting samples from six excavation sites along the entire length of the Nile valley spanning 4000 years of Egyptian history. Samples from 17 mummies and 14 skeletal remains were collected, and high quality mitochondrial genomes were reconstructed from 10 individuals. According to the authors the analyzed mitochondrial genomes matched the results from the 2017 study at Abusir el-Meleq.[18]
A 2020 study was conducted on ancient samples from Lebanon. Two individuals who lived in Lebanon around 500 BCE did not cluster with their contemporary Lebanese population. The study used the same Egyptian samples from the 2017 Schuenemann et al. study to further test these two individuals. One of these two individuals was a female who formed a clad with the three ancient Egyptian individuals from Schuenemann et al., implying that she shared all of her ancestry with them or a genetically equivalent population. The other one was a male who derived ~70% of his ancestry from a population related to the female and ~30% from a population related to ancient Levantines. Further testing suggests that the female was an Egyptian woman and the male was her son from a man who himself had both Egyptian and Lebanese ancestries.[19]
2018 study of Nakht-Ankh and Khnum-Nakht
The tomb of two high-status Egyptians, Nakht-Ankh and Khnum-Nakht, was discovered by Egyptian workmen directed by Sir William Flinders Petrie and Ernest Mackay in 1907. Nakht-Ankh and Khnum-Nakht lived during the 12th Dynasty (1985–1773 BCE) in Middle Egypt and were aged 20 years apart. Their tomb has been called Tomb of Two Brothers because the mummies were buried adjacent to one other and inscriptions on the coffins mention the female name Khnum-Aa, who is described as 'lady of the house' and referred to as the mother of both Nakht-Ankh and Khnum-Nakht. The Y-chromosome sequences were not complete, but the Y-chromosome SNPs indicated that they had different fathers, suggesting that they were half-brothers. The SNP identities were consistent with mtDNA haplogroup M1a1 with 88.05–91.27% degree of confidence, thus "confirming the African origins of the two individuals" according to the study authors, based on their maternal lineage.[20][21] According to the results of analysis published by FTDNA in 2023, Nakht Ankh's most likely Y-DNA haplogroup was H-Z19008, a subclade of H2.[22]
2018 study of Djehutynakht
In 2018 the mummified head of Djehutynakht was analysed for mitochondrial DNA. Djehutynakht was the nomarch of the Hare nome in Upper Egypt during the 11th or 12th Dynasty in the early Middle Kingdom period, c. 2000 BC. Two laboratories independently analysed Djehutynakht's DNA and found that he belonged to the mtDNA haplogroup U5b2b5.[23] Related sequences have been observed in ancient DNA from Europe and Phoenicia, with no direct matches to the Djehutynakht sequence.[23] Among ancient Egyptian samples the Djehutynakht sequence resembles a U5a lineage from sample JK2903, a much more recent 2000-year-old skeleton from the Abusir el-Meleq site in Egypt.[9] Haplogroup U5 is found in modern Egyptians,[24][25] and is found in modern Egyptian Berbers from the Siwa Oasis in Egypt. A 2009 study by Coudray et al. recorded haplogroup U5 at 16.7% in the Siwa Oasis in Egypt, whereas haplogroup U6 is more common in other Berber populations to the west of Egypt.[26]
2020 study on two Egyptian child mummies
A study on male child mummies from the Greco-Roman period originating in the Memphite or Luxor area, revealed that the mtDNA for one was T2c1a and the other HV. Identical or phylogenetically close derivatives of these lineages are present in both ancient and modern Egyptians, as well as among several present-day populations of the Near East and North Africa. The researchers noted that mtDNA alone is not enough to reach any precise conclusion about the origin of an individual, but the results are in accordance with an Egyptian origin. The ages of the two mummified corpses ranged from 11-15 years old, and 2-4 years old.[27]
2020 study of Tutankhamun and other mummies of the 18th Dynasty
A 2020 study by Gad, Hawass, et al. analysed mitochondrial and Y-chromosomal haplogroups from Tutankhamun's family members of the 18th Dynasty, using comprehensive control procedures to ensure quality results. The study found that the Y-chromosome haplogroup of the family was R1b.[28][29] Haplogroup R1b is carried by modern Egyptians.[28] Modern Egypt is also the only African country that is known to harbor all three R1 subtypes, including R1b-M269.[30] The Y-chromosome profiles for Tutankhamun and Amenhotep III were incomplete and the analysis produced differing probability figures despite having concordant allele results. Because the relationships of these two mummies with the KV55 mummy (identified as Akhenaten) had previously been confirmed in an earlier study, the haplogroup prediction of both mummies could be derived from the full profile of the KV55 data.[28] Genetic analysis indicated the following haplogroups for the 18th Dynasty:
- Amenhotep III: YDNA R1b & mtDNA H2b.
- Tutankhamun: YDNA R1b & mtDNA K.
- Akhenaten: YDNA R1b & mtDNA K.
- Tiye: mtDNA K.
- Yuya: YDNA G2a & mtDNA K.
- Thuya: mtDNA K.
Both Y-DNA haplogroups R1b and G2a, as well as both mtDNA haplogroups H and K, are carried by modern Egyptians.[28]
In a comment on Hawas et al. (2010[31]&2012[8]), Keita pointed out, based on inserting the data into the PopAffiliator online calculator, which only calculates affinity to East Asia, Eurasia, and sub-Saharan Africa, but not to North Africa or the Near East, for instance,[32] that the majority of the samples: "have an affinity with sub-Saharan Africans in one affinity analysis, which does not mean that they lacked other affiliations—an important point that typological thinking obscures. Also, different data and algorithms might give different results, which would illustrate the complexity of biological heritage and its interpretation."[33]
2020 study of Takabuti
In 2020 the mummy of Takabuti was tested for mitochondrial DNA. Takabuti was a noblewoman from Thebes in Upper Egypt who lived during the 25th Dynasty, c. 660 BC. Analysis of her DNA revealed that she belonged to the mtDNA haplogroup H4a1.[34] The study states that “the H super-haplogroup is the most common mtDNA lineage in Europe and is found also in parts of present-day Africa and western Asia”.[34] Haplogroup H is also carried by modern Egyptians, the subgroup H4 in particular, along with H2a1 and H13a1, account for 42% of H lineages in modern Egypt, with H4 accounting for 16.7% of H lineages in modern Egypt.[35]
2020 study of mummies at the Kurchatov Institute
In 2020, three mummies, dating from the 1st millennium BCE, from the Pushkin Museum of Arts collection were tested at the Kurchatov Institute of Moscow for their mitochondrial and Y-chromosomal haplogroups. Two of the mummies were found to belong to the Y-chromosomal haplogroup R1b1a1b (R1b-M269),[36] which originated either in Eastern Europe or in the Near East, and to the Y-chromosome haplogroup E1b1b1a1b2a4b5a,[36] which originated either in Southeastern Europe or in the Near East, in the early Holocene after a Paleolithic exit out of Africa, specifically out of Egypt, of E-M78 chromosomes devoid of the V13 mutation.[37][38] They also belonged to mtDNA haplogroups L3h1 and N5, common in Northern Africans and Middle Easterners, respectively. The third mummy was found to belong to mtDNA haplogroup N, widely distributed throughout Eurasia.[36]
Genetic studies on modern Egyptians
Genetic analysis of modern Egyptians reveals that they have paternal lineages common to other indigenous Afroasiatic-speaking populations in North Africa, West Asia, Anatolia and Horn of Africa; Some studies have proposed the view that these lineages would have spread into North Africa and Horn of Africa from Western Asia during the Neolithic Revolution and were maintained by the predynastic period.[39][40]
A study by Krings et al. (1999) on mitochondrial DNA clines along the Nile Valley found that a Eurasian cline runs from Northern Egypt to Southern Sudan and a Sub-Saharan cline from Southern Sudan to Northern Egypt, derived from a sample size of 224 individuals (68 Egyptians, 80 Nubians, 76 southern Sudanese). The study also found Egypt and Nubia have low and similar amounts of divergence for both mtDNA types, which is consistent with historical evidence for long-term interactions between Egypt and Nubia. However, there are significant differences between the composition of the mtDNA gene pool of the Egyptian samples and that of the Nubians and Southern Sudanese samples. The diversity of the Eurasian mtDNA type was highest in Egypt and lowest in southern Sudan, whereas the diversity of the sub-Saharan mtDNA type was lowest in Egypt and highest in southern Sudan. The authors suggested in their conclusion that Egypt and Nubia had more genetic contact than either did with southern Sudan and that the migration from north to south was either earlier or lesser in the extent of gene flow than the migration from south to north.[41]
A study by Luis et al. (2004) found that the male haplogroups in a sample of 147 Egyptians were E1b1b (36.1%, predominantly E-M78), J (32.0%), G (8.8%), T (8.2%), and R (7.5%).[1] The study found that "Egypt's NRY frequency distributions appear to be much more similar to those of the Middle East than to any sub-Saharan African population, suggesting a much larger Eurasian genetic component ... The cumulative frequency of typical sub-Saharan lineages (A, B, E1b1a) is 3.4% in Egypt ... whereas the haplogroups of Eurasian origin (Groups C, D, and F–Q) account for 59% [in Egypt]."[1] E1b1b subclades are characteristic of some Afro-Asiatic speakers and are believed to have originated in either the Middle East, North Africa, or the Horn of Africa.[1][42][43] Cruciani et al. (2007) suggests that E-M78, E1b1b predominant subclade in Egypt, originated in Northeastern Africa (Egypt and Libya in the study), with a corridor for bidirectional migrations between northeastern and eastern Africa (at least 2 episodes between 23.9 and 17.3 ky and 18.0–5.9 ky ago), trans-Mediterranean migrations directly from northern Africa to Europe (mainly in the last 13.0 ky), and flow from northeastern Africa to western Asia between 20.0 and 6.8 ky ago. Cruciani et al. proposed that E-M35, the parent clade of E-M78, originated in Eastern Africa during the Palaeolithic and subsequently spread to Northeastern Africa, 23.9–17.3 ky ago. Cruciani et al. also state that the presence of E-M78 chromosomes in Eastern Africa can be only explained through a back migration of chromosomes that had acquired the M78 mutation in Northeast Africa.[43]
Other studies have shown that modern Egyptians have genetic affinities primarily with populations of North Africa and the Middle East,[40][39] and to a lesser extent the Horn of Africa and European populations.[44][45] Another study states that "the information available on individual groups in Ethiopia and North Africa is fairly limited but sufficient to show that they are all separate from sub-Saharan Africans and that North Africans and East Africans (such as Ethiopians) are clearly separate", and concluded that most Ethiopians came from an admixture and that the larger fraction of Sub-Saharan genes came during the Neolithic times "before the beginning of the Egyptian civilisation".[46] The study also found the gene frequency of North African populations and, to a lesser extent, East Africa to be intermediate between Africa and Europe.[47] In addition, some studies suggest ties with populations in the Middle East, as well as some groups in southern Europe,[39] and a closer link to other North Africans.[40]
A 2004 mtDNA study of 58 upper Egyptian individuals included 34 individuals from Gurna, a small settlement on the hills opposite Luxor. The 34 individuals from Gurna exhibited the haplogroups: M1 (6/34 individuals, 17.6%), H (5/34 individuals, 14.7%), L1a (4/34 individuals, 11.8%) and U (3/34 individuals, 8.8%). The M1 haplotype frequency in Gurna individuals (6/34 individuals, 17.6%) is similar to that seen in Ethiopian population (20%), along with a West Eurasian component different in haplogroup distribution in the Gurna individuals. However, the M1 haplotypes from Gurna individuals exhibited a mutation that is not present in Ethiopian population; whereas this mutation was present in non-M1 haplotype individuals from Gurna. Nile Valley Egyptians do not show the characteristics that were shown by the Gurna individuals. The results of the study suggested that the sample of Gurna individuals had retained elements of an ancestral genetic structure from an ancestral East African population, characterized by a high M1 haplogroup frequency.[45] Another 2004 mtDNA study featured the Gurna individuals samples, and clustered them together with the Ethiopian and Yemeni groups, in between the Near Eastern and other African sample groups.[48]
A 2005 genetic study found close affinities of eastern sub-Saharan populations with Egypt in the phylogenetic trees through analysis of the short DNA sequences. The authors suggested that the influential role of the Nile River served as a migratory route and an agent of genetic flow which contributed to present-day heterogeneity in Egypt.[49]
Though there has been much debate of the origins of haplogroup M1 a 2007 study had concluded that M1 has West Asia origins not a Sub Saharan African origin, although the majority of the M1a lineages found outside and inside Africa had a more recent East African origin, as a result of "the first M1 backflow [from Asia] to Africa, dated around 30,000 [years ago]". The study states that "the most ancient dispersals of M1 occurred in northwestern Africa, reaching also the Iberian Peninsula, instead of Ethiopia", and states that the evidence points to either "that the Near East was the most probable origin of the primitive M1 dispersals, West into Africa and East to Central Asia ... [with] the Sinai Peninsula as the most probable gate of entrance of this backflow to Africa" or "that M1 is an autochthonous North African clad that had its earliest spread in northwestern areas marginally reaching the Near East and beyond".[50]
However, other authors have proposed the view that the M haplogroup developed in Africa before the 'Out of Africa' event around 50,000 years ago, and dispersed from North Africa or East Africa 10,000 to 20,000 years ago. Quintana-Murci et al. stated in reference to the M haplogroup that "Its variation and geographical distribution suggest that Asian haplogroup M separated from eastern-African haplogroup M more than 50,000 years ago. Two other variants (489C and 10873C) also support a single origin of haplogroup M in Africa".[51][52]
A 2003 Y-chromosome study was performed by Lucotte on modern Egyptians, with haplotypes V, XI, and IV being most common. Haplotype V is common among all North Africans and has a low frequency outside the North African region. Haplotypes V, XI, and IV are all predominantly North African/Horn of African haplotypes, and they are far more dominant in Egyptians than in Middle Eastern or European groups.[53] The pattern of diversity for these variants in the Egyptian Nile Valley was largely the product of population events that occurred in the late Pleistocene to mid-Holocene through the First Dynasty.[53]
Keita (2008) examined a published Y-chromosome dataset on Afro-Asiatic populations and remarked that a key subclade of E-M35, namely the key lineage E-M78, was shared between the populations in the locale of original Egyptian speakers and modern Cushitic speakers from the Horn of Africa. These lineages are present in modern Egyptians, Berbers, Cushitic speakers from the Horn of Africa, and Semitic speakers in the Near-East. He noted that variants are also found in the Aegean and Balkans. The origin of E-M35 was in East Africa, whereas the origin of its subclade mutation E-M78 was in Northeast Africa (the area of Egypt and Libya).[54] This parent clade and its subclade mutations in general were dominant in a core portion of Afro-Asiatic speaking populations which included Cushitic, Egyptian, Berber groups, and Semitic speakers from the Near East, where Semitic speakers showed a decline in frequency going west to east in the Levantine-Syria region. He concluded that "the genetic data give population profiles that clearly indicate males of African origin, as opposed to being of Asian or European descent" but acknowledged that the biodiversity does not indicate any specific set of skin colors or facial features as populations were subject to microevolutionary pressures.[55]
Babiker, H et al. (2011) examined the genotypes of 15 STRs for 498 individuals from 18 Sudanese populations and featured comparative genotype data with Egypt, Somalia and the Karamoja population from Uganda. Overall, the results showed that the genotypes of individuals from northern Sudan clustered with those of Egypt, the Somali population was found to be genetically distinct and individuals from southern Sudan clustered with those from the Karamoja population. The study determined that similarity of the Nubian and Egyptian populations suggested that migration, potentially bidirectional, occurred along the Nile river Valley, which is consistent with the historical evidence for long-term interactions between Egypt and Nubia.[56]
A study by Hollfelder et al. (2017) analyzed various populations and found that Copts and Egyptians showed low levels of genetic differentiation and lower levels of genetic diversity compared to the northeast African groups. Copts and Egyptians displayed similar levels of European/Middle Eastern ancestry (Copts were estimated to be of 69.54% ± 2.57 European ancestry, and the Egyptians of 70.65% ± 2.47 European ancestry). The study concluded that the Copts and the Egyptians have a common history linked to smaller population sizes. The behavior in the admixture analyses is consistent with shared ancestry between Copts and Egyptians and/or additional genetic drift in the Copts.[57] An allele frequency comparative study conducted in 2020 between the two main Egyptian ethnic groups, Muslims and Christians, each group represented by a sample of 100 unrelated healthy individuals, supported the conclusion that Egyptian Muslims and Egyptian Christians genetically originate from the same ancestors.[58]
Y-DNA haplogroups
A study by Arredi et al., which analyzed 275 samples from five populations in Algeria, Tunisia, and Egypt, as well as published data from Moroccan populations, suggested that the North African pattern of Y-chromosomal variation, including in Egypt, is largely of Neolithic origin. The study analyzed North African populations, including North Egyptians and South Egyptians, as well as samples from southern Europe, the Middle East, and sub-Saharan Africa, and revealed the following conclusions about the male-lineage variation in North Africa: "The lineages that are most prevalent in North Africa are distinct from those in the regions to the immediate north and south: Europe and sub-Saharan Africa ... two haplogroups predominate within North Africa, together making up almost two-thirds of the male lineages: E3b2 and J* (42% and 20%, respectively). E3b2 is rare outside North Africa, and is otherwise known only from Mali, Niger, and Sudan to the immediate south, and the Near East and Southern Europe at very low frequencies. Haplogroup J reaches its highest frequencies in the Middle East".[40]
A study by Lucotte using the Y-chromosome of 274 male individuals (162 from Lower Egypt, 66 from Upper Egypt , 46 from Lower Nubia) found that the main haplotype V has higher frequency in the North than in the South, and haplotype XI has higher frequency in the South than in the North, whereas haplotype IV is found in the South (highest in Lower Nubia). The study states that haplotype IV is also characteristic of Sub-Saharan populations.[59] Remarking on Lucotte's Y-chromosome study, which found that haplotypes V, XI, and IV are most common, Keita states that "a synthesis of evidence from archaeology, historical linguistics, texts, distribution of haplotypes outside Egypt, and some demographic considerations lends greater support to the establishment, before the Middle Kingdom, of the observed distributions of the most prevalent haplotypes V, XI, and IV. It is suggested that the pattern of diversity for these variants in the Egyptian Nile Valley was largely the product of population events that occurred in the late Pleistocene to mid-Holocene through the First Dynasty".[53] Keita later states "Later, mid-Holocene climatic-driven migrations led to a major settlement of the valley in Upper Egypt and Nubia, but less so in Lower Egypt, by diverse Saharans with haplotypes IV, XI, and V. These people fused with the indigenous valley peoples, as did Near Easterners with VII and VIII, but perhaps also some V".[60]
The major downstream mutations within the M35 subclade are M78 and M81. There are also other M35 lineages, e.g., M123. In Egypt, haplotypes VII and VIII are associated with the J haplogroup, which is predominant in the Near East.[53]
| Population | Nb | A/B | E1b1a | E1b1b1
(M35) |
E1b1b1a
(M78) |
E1b1b1b1
(M81) |
E1b1b1b2
(M123,M34) |
F | K | G | I | J1 | J2 | R1a | R1b | Other | Study |
| Egyptians | 110 | 0 | 3.5% | 0 | 36% | 0 | 8.5% | 0 | 0 | 7.5% | 0 | 24.5% | 2% | 2.8% | 8.4% | 6.5% | Fadhloui-Zid et al. (2013) |
| Egyptians | 370 | 1.35% | 2.43% | 3.24% | 21.89% | 11.89% | 6.76% | 1.08% | 0.27% | 5.68% | 0.54% | 20.81% | 6.75% | 2.16% | 5.94% | 9.21% | Bekada et al. (2013)[35] |
| Egyptians | 147 | 2.7% | 2.7% | 0 | 18.4% | 5.4% | 0 | 0 | 8.2% | 8.8% | 0 | 19.7% | 12.2% | 3.4% | 4.1% | 2.1% | Luis et al. (2004)[1] |
| Egyptians from El-Hayez Oasis (Western Desert) | 35 | 0 | 5.70% | 5.7% | 28.6% | 28.6% | 0 | 0 | 0 | 0 | 0 | 31.4% | 0 | 0 | 0 | 0 | Kujanová et al. (2009)[10] |
| Berbers from Siwa Oasis (Western Desert) | 93 | 28.0% | 6.5% | 2.2% | 6.5% | 1% | 0 | 0 | 0 | 3.2% | 0 | 7.5% | 6.5% | 0 | 28.0% | 8.3% | Dugoujon et al. (2009)[61] |
| Egyptians | 87 | 1% | 3% | 10% | 31% | 0 | 2.5% | 0 | 0 | 2% | 0 | 20% | 15% | 5% | 2% | 8.5% | Pagani et al. (2015) |
| Northern Egyptians | 44 | 2.3% | 0 | 4.5% | 27.3% | 11.3% | 0 | 6.8% | 2.3% | 0 | 0 | 9.1% | 9.1% | 2.3% | 9.9% | 6.8% | Arredi et al. (2004) |
| Southern Egyptians | 29 | 0.0% | 0 | 0 | 17.2% | 6.8% | 0 | 17.2% | 10.3% | 0 | 3.4% | 20.7% | 3.4% | 0 | 13.8% | 0 | Arredi et al. (2004) |
- Distribution of E1b1b1a (E-M78) and its subclades
| Population | N | E-M78 | E-M78* | E-V12* | E-V13 | E-V22 | E-V32 | E-V65 | Study |
| Egyptians (sample includes people labeled as "berber" and people from the oases) | 370 | 21.89% | 0.81% | 7.03% | 0.81% | 9.19% | 1.62% | 2.43% | Bekada et al. (2013)[35] |
| Southern Egyptians | 79 | 50.6% | 44.3% | 1.3% | 3.8% | 1.3% | Cruciani et al. (2007)[3] | ||
| Egyptians from Bahari | 41 | 41.4% | 14.6% | 2.4% | 21.9% | 2.4% | Cruciani et al. (2007) | ||
| Northern Egyptians (Delta) | 72 | 23.6% | 5.6% | 1.4% | 13.9% | 2.8% | Cruciani et al. (2007) | ||
| Egyptians from Gurna Oasis | 34 | 17.6% | 5.9% | 8.8% | 2.9% | Cruciani et al. (2007) | |||
| Egyptian from Siwa Oasis | 93 | 6.4% | 2.1% | 4.3% | Cruciani et al. (2007) |
Mitochondrial DNA
In 2009 mitochondrial data was sequenced for 277 unrelated Egyptian individuals[62] by Jessica L Saunier et al. in the journal Forensic Science International, as follows
- R0 and its subgroups (31.4%)
- L3 (12.3%); and Asian origin (n = 33)
including M (6.9%)
- T (9.4%)
- U (9.0%)
- J (7.6%)
- N (5.1%)
- K (4.7%)
- L2 (3.6%)
- L1 (2.5%)
- I (3.2%)
- W (0.7%)
- X (1.4%); African origin (n = 57) including L0 (2.2%)
Autosomal DNA
Mohamed, T et al. (2009) in their study of nomadic Bedouins featured a comparative study with a worldwide population database and a sample size of 153 Bedouin males. Their analysis discovered that both Muslim Egyptians and Coptic Christians showed a distinct North African cluster at 65%. This is their predominant ancestral component, and unique to the geographic region of Egypt.[63]
In a 2019 study that analyzed the autosomal make-up of 21 modern North African genomes and other populations using Ancient DNA reference populations, this sample of Egyptian genomes were found to share more affinity with Middle Eastern populations compared to other North Africans. Egyptians carry more of the Caucasus hunter gatherer / Iran Neolithic component compared to other North Africans, more of the Natufian related component and less of the Iberomaurusian related component than other North Africans, and also less of the Steppe / European hunter gatherer component, consistent with Egypt's geographical proximity to southwest Asia.[64]
Coptic Christians
A Y-DNA sample of Copts from Egypt was analyzed in Éric Crubézy et al. 2010. The Y-DNA profile was:
- 74% E1b1
- 7% G
- 3% T
- 1% J1
- 15% ambiguous.[65]
Cruzby further added that “The haplotype E1b1 has a fairly wide distribution across Africa, but never had been described with such frequency in this area”. He also noted that Ethiopia was one of the areas in the world where the haplotype was most common. Cruzby suggested that this shared lineage could be attributed to the contacts between Coptic Christian communities in both Egypt and Ethiopia although the overlaps between the religious communities were rare.[66]
An allele frequency comparative study conducted in 2020 between the two main Egyptian ethnic groups, Muslims and Christians, each group represented by a sample of 100 unrelated healthy individuals, supported the conclusion that Egyptian Muslims and Egyptian Christians genetically originate from the same ancestors.[58]
In Sudan
According to Y-DNA analysis by Hassan et al. (2008), 45% of Copts in Sudan (of a sample of 33) carry haplogroup J1. Next most common was E1b1b, the most common haplogroup in North Africa. Both paternal lineages are common among other regional Afroasiatic-speaking populations, such as Beja, Ethiopians, and Sudanese Arabs, as well as non-Afroasiatic-speaking Nubians.[67] E1b1b reaches its highest frequencies among North African and Horn of Africa populations such as Amazighs and Somalis.[68] The next most common haplogroups borne by Copts are R1b (15%), most common in Europe, and the widespread African haplogroup B (15%). According to the study, the presence of haplogroup B may also be consistent with the historical record in which southern Egypt was colonized by Nilotic populations during the early state formation.[67]
Maternally, Hassan (2009) found that the majority of Copts in Sudan (of a sample of 29) carried descendants of the macrohaplogroup N; of these, haplogroup U6 was most frequent (28%), followed by T1 (17%). In addition, Copts carried 14% M1 and 7% L1c.[69]
A 2015 study by Dobon et al. identified an ancestral autosomal component of West Eurasian origin that is common to many modern Afroasiatic-speaking populations in Northeast Africa. Known as the Coptic component, it peaks among Egyptian Copts who settled in Sudan over the past two centuries, they also formed a separated group in PCA, a close outlier to other Egyptians, Afroasiatic-speaking Northeast Africans and Middle East populations. The Coptic component evolved out of a main North African and Middle Eastern ancestral component that is shared by other Egyptians and also found at high frequencies among other Afroasiatic-speaking populations in Northeast Africa (~70%), who carry a Nilo-Saharan element as well. The scientists suggest that this points to a common origin for the general population of Egypt.[70] They also associate the Coptic component with Ancient Egyptian ancestry, without the later Arabic influence that is present among other Egyptians, especially people of the Sinai.[70]
In another 2017 study that genotyped and analyzed the same populations including Sudanese Copts and Egyptians, The ADMIXTURE analyses and the PCA displayed the genetic affinity of the Copts to the Egyptian population. Assuming few clusters, the Copts appeared admixed between Near Eastern/European populations and northeastern Sudanese and look similar in their genetic profile to the Egyptians. Assuming greater number of clusters (K≥18), the Copts formed their own separate ancestry component that was shared with Egyptians but can also be found in Arab populations. This behavior in the admixture analyses is consistent with shared ancestry between Copts and Egyptians and/or additional genetic drift in the Copts. The Egyptians and Copts showed low levels of genetic differentiation (FST = 0.00236), lower levels of genetic diversity and greater levels of RoH compared to other northeast African groups, including Arab and Middle Eastern groups that share ancestry with the Copts and Egyptians. A formal test did not find significant admixture into the Egyptians from other tested groups (X), and the Copts and Egyptians displayed similar levels of European or Middle Eastern ancestry (Copts were estimated to be of 69.54% ± 2.57 European ancestry, and the Egyptians of 70.65% ± 2.47 European ancestry). Taken together, these results point to that the Copts and the Egyptians have a common history linked to smaller population sizes, and that Sudanese Copts have remained relatively isolated since their arrival to Sudan with only low levels of admixture with local northeastern Sudanese groups.[57]
See also
References
- 1 2 3 4 5 Luis JR, Rowold DJ, Regueiro M, Caeiro B, Cinnioğlu C, Roseman C, Underhill PA, Cavalli-Sforza LL, Herrera RJ (March 2004). "The Levant versus the Horn of Africa: evidence for bidirectional corridors of human migrations". American Journal of Human Genetics. 74 (3): 532–44. doi:10.1086/382286. PMC 1182266. PMID 14973781.
- ↑ Krause, Johannes; Schiffels, Stephan (30 May 2017). "Ancient Egyptian mummy genomes suggest an increase of Sub-Saharan African ancestry in post-Roman periods". Nature Communications. 8: 15694. Bibcode:2017NatCo...815694S. doi:10.1038/ncomms15694. PMC 5459999. PMID 28556824.
- 1 2 3 Cruciani F, La Fratta R, Trombetta B, Santolamazza P, Sellitto D, Colomb EB, Dugoujon JM, Crivellaro F, Benincasa T, Pascone R, Moral P, Watson E, Melegh B, Barbujani G, Fuselli S, Vona G, Zagradisnik B, Assum G, Brdicka R, Kozlov AI, Efremov GD, Coppa A, Novelletto A, Scozzari R (June 2007). "Tracing past human male movements in northern/eastern Africa and western Eurasia: new clues from Y-chromosomal haplogroups E-M78 and J-M12". Molecular Biology and Evolution. 24 (6): 1300–11. doi:10.1093/molbev/msm049. PMID 17351267.
- ↑ (PDF). 26 June 2008 https://web.archive.org/web/20080626010050/http://www.familytreedna.com/pdf/hape3b.pdf. Archived from the original (PDF) on 26 June 2008. Retrieved 13 April 2023.
{{cite web}}: Missing or empty|title=(help) - ↑ Wood, Elizabeth T.; et al. (2005). "Contrasting patterns of Y chromosome and mtDNA variation in Africa: evidence for sex-biased demographic processes" (PDF). European Journal of Human Genetics. 13 (7): 867–876. doi:10.1038/sj.ejhg.5201408. PMID 15856073. S2CID 20279122. Archived (PDF) from the original on 24 September 2016. Retrieved 24 September 2016.
- ↑ Kemp, Barry J. (7 May 2007). Ancient Egypt: Anatomy of a Civilisation. Routledge. pp. 46–58. ISBN 9781134563883. Archived from the original on 14 April 2023. Retrieved 14 April 2023.
- ↑ Jr, William H. Stiebing; Helft, Susan N. (3 July 2023). Ancient Near Eastern History and Culture. Taylor & Francis. pp. 209–212. ISBN 978-1-000-88066-3.
- 1 2 Hawass, Zahi; et al. (2012). "Revisiting the harem conspiracy and death of Ramesses III: anthropological, forensic, radiological, and genetic study". BMJ. 345 (e8268): e8268. doi:10.1136/bmj.e8268. hdl:10072/62081. PMID 23247979. S2CID 206896841.
- 1 2 3 4 5 6 7 8 9 10 11 12 Krause, Johannes; Schiffels, Stephan (30 May 2017). "Ancient Egyptian mummy genomes suggest an increase of Sub-Saharan African ancestry in post-Roman periods". Nature Communications. 8: 15694. Bibcode:2017NatCo...815694S. doi:10.1038/ncomms15694. PMC 5459999. PMID 28556824.
- 1 2 Kujanová M, Pereira L, Fernandes V, Pereira JB, Cerný V (October 2009). "Near eastern neolithic genetic input in a small oasis of the Egyptian Western Desert". American Journal of Physical Anthropology. 140 (2): 336–46. doi:10.1002/ajpa.21078. PMID 19425100.
- ↑ Pagani, Luca; Schiffels, Stephan; Gurdasani, Deepti; Danecek, Petr; Scally, Aylwyn; Chen, Yuan; Xue, Yali; Haber, Marc; Ekong, Rosemary; Oljira, Tamiru; Mekonnen, Ephrem (4 June 2015). "Tracing the route of modern humans out of Africa by using 225 human genome sequences from Ethiopians and Egyptians". American Journal of Human Genetics. 96 (6): 986–991. doi:10.1016/j.ajhg.2015.04.019. ISSN 1537-6605. PMC 4457944. PMID 26027499.
- ↑ Eltis, David; Bradley, Keith R.; Perry, Craig; Engerman, Stanley L.; Cartledge, Paul; Richardson, David (12 August 2021). The Cambridge World History of Slavery: Volume 2, AD 500-AD 1420. Cambridge University Press. p. 150. ISBN 978-0-521-84067-5.
- ↑ Keita Shomarka. (2022). "Ancient Egyptian "Origins and "Identity" In Ancient Egyptian society : challenging assumptions, exploring approaches. Abingdon, Oxon. pp. 111–122. ISBN 978-0367434632.
{{cite book}}: CS1 maint: location missing publisher (link) - ↑ Candelora, Danielle (31 August 2022). Candelora, Danielle; Ben-Marzouk, Nadia; Cooney, Kathyln (eds.). Ancient Egyptian society : challenging assumptions, exploring approaches. Abingdon, Oxon. pp. 101–111. ISBN 9780367434632.
{{cite book}}: CS1 maint: location missing publisher (link) - ↑ Jr, William H. Stiebing; Helft, Susan N. (3 July 2023). Ancient Near Eastern History and Culture. Taylor & Francis. pp. 209–212. ISBN 978-1-000-88066-3.
- ↑ Ehret, Christopher (20 June 2023). Ancient Africa: A Global History, to 300 CE. Princeton: Princeton University Press. pp. 83–86, 167–169. ISBN 978-0-691-24409-9. Archived from the original on 22 March 2023. Retrieved 20 March 2023.
- ↑ Ehret, Christopher (20 June 2023). Ancient Africa: A Global History, to 300 CE. Princeton University Press. p. 167. ISBN 978-0-691-24410-5.
- ↑ "Human mitochondrial haplogroups and ancient DNA preservation across Egyptian history (Urban et al. 2021)" (PDF). ISBA9, 9th International Symposium on Biomolecular Archaeology, p.126. 2021. Archived (PDF) from the original on 16 October 2021. Retrieved 14 January 2023.
In a previous study, we assessed the genetic history of a single site: Abusir el-Meleq from 1388 BCE to 426 CE. We now focus on widening the geographic scope to give a general overview of the population genetic background, focusing on mitochondrial haplogroups present among the whole Egyptian Nile River Valley. We collected 81 tooth, hair, bone, and soft tissue samples from 14 mummies and 17 skeletal remains. The samples span approximately 4000 years of Egyptian history and originate from six different excavation sites covering the whole length of the Egyptian Nile River Valley. NGS 127 based ancient DNA 8 were applied to reconstruct 18 high-quality mitochondrial genomes from 10 different individuals. The determined mitochondrial haplogroups match the results from our Abusir el-Meleq study.
- ↑ Haber, Marc; Nassar, Joyce; Almarri, Mohamed A.; Saupe, Tina; Saag, Lehti; Griffith, Samuel J.; Doumet-Serhal, Claude; Chanteau, Julien; Saghieh-Beydoun, Muntaha; Xue, Yali; Scheib, Christiana L.; Tyler-Smith, Chris (2 July 2020). "A Genetic History of the Near East from an aDNA Time Course Sampling Eight Points in the Past 4,000 Years". American Journal of Human Genetics. 107 (1): 149–157. doi:10.1016/j.ajhg.2020.05.008. PMC 7332655. PMID 32470374.
- ↑ Konstantina; Drosoua Campbell Price; Terence A. Brown (February 2018). "The kinship of two 12th Dynasty mummies revealed by ancient DNA sequencing". Journal of Archaeological Science: Reports. 17: 793–797. Bibcode:2018JArSR..17..793D. doi:10.1016/j.jasrep.2017.12.025. Archived from the original on 17 January 2022. Retrieved 12 November 2021.
- ↑ "The kinship of two 12th Dynasty mummies revealed by ancient DNA sequencing" (PDF).
- ↑ "FamilyTreeDNA - Notable Connections - Nakht-Ankh". FamilyTreeDNA Discover (Beta). Retrieved 15 September 2023.
- 1 2 Loreille, O.; Ratnayake, S.; Bazinet, A. L.; Stockwell, T. B.; Sommer, D. D.; Rohland, N.; Mallick, S.; Johnson, P. L.; Skoglund, P.; Onorato, A. J.; Bergman, N. H.; Reich, D.; Irwin, J. A. (2018). "Biological Sexing of a 4000-Year-Old Egyptian Mummy Head to Assess the Potential of Nuclear DNA Recovery from the Most Damaged and Limited Forensic Specimens". Genes. 9 (3): 135. doi:10.3390/genes9030135. PMC 5867856. PMID 29494531.
- ↑ Elmadawy, Mostafa Ali; Nagai, Atsushi; Gomaa, Ghada M.; Hegazy, Hanaa M. R.; Shaaban, Fawzy Eid; Bunai, Yasuo (2013). "Investigation of mtDNA control region sequences in an Egyptian population sample". Legal Medicine (Tokyo, Japan). 15 (6): 338–341. doi:10.1016/j.legalmed.2013.06.006. ISSN 1873-4162. PMID 23910099.
- ↑ Saunier, Jessica L.; Irwin, Jodi A.; Strouss, Katharine M.; Ragab, Hisham; Sturk, Kimberly A.; Parsons, Thomas J. (2009). "Mitochondrial control region sequences from an Egyptian population sample". Forensic Science International. Genetics. 3 (3): e97–103. doi:10.1016/j.fsigen.2008.09.004. ISSN 1878-0326. PMID 19414160.
- ↑ Coudray C, Olivieri A, Achilli A, Pala M, Melhaoui M, Cherkaoui M, et al. (March 2009). "The complex and diversified mitochondrial gene pool of Berber populations". Annals of Human Genetics. 73 (2): 196–214. doi:10.1111/j.1469-1809.2008.00493.x. PMID 19053990. S2CID 21826485.
- ↑ Oras, Ester; Anderson, Jaanika; Tõrv, Mari; Vahur, Signe; Rammo, Riina; Remmer, Sünne; Mölder, Maarja; Malve, Martin; Saag, Lehti; Saage, Ragnar; Teearu-Ojakäär, Anu; Peets, Pilleriin; Tambets, Kristiina; Metspalu, Mait; Lees, David C. (2020). "Multidisciplinary investigation of two Egyptian child mummies curated at the University of Tartu Art Museum, Estonia (Late/Graeco-Roman Periods)". PLOS ONE. 15 (1): e0227446. Bibcode:2020PLoSO..1527446O. doi:10.1371/journal.pone.0227446. ISSN 1932-6203. PMC 6964855. PMID 31945091.
- 1 2 3 4 Gad, Yehia (2020). "Maternal and paternal lineages in King Tutankhamun's family". Guardian of Ancient Egypt: Essays in Honor of Zahi Hawass. Czech Institute of Egyptology. pp. 497–518. ISBN 978-80-7308-979-5.
- ↑ Gad, Yehia (2020). "Insights from ancient DNA analysis of Egyptian human mummies: clues to disease and kinship". Human Molecular Genetics. 30 (R1): R24–R28. doi:10.1093/hmg/ddaa223. PMID 33059357. Archived from the original on 2 May 2021. Retrieved 19 December 2022.
- ↑ Luis JR, Rowold DJ, Regueiro M, Caeiro B, Cinnioğlu C, Roseman C, Underhill PA, Cavalli-Sforza LL, Herrera RJ (March 2004). "The Levant versus the Horn of Africa: evidence for bidirectional corridors of human migrations". American Journal of Human Genetics. 74 (3): 532–44. doi:10.1086/382286. PMC 1182266. PMID 14973781.
- ↑ Hawass, Zahi (2010). "Ancestry and pathology in King Tutankhamun's family". JAMA. 303 (7): 638–647. doi:10.1001/jama.2010.121. PMID 20159872. Archived from the original on 14 April 2020. Retrieved 21 March 2023.
- ↑ Pereira, Luísa; Alshamali, Farida; Andreassen, Rune; Ballard, Ruth; Chantratita, Wasun; Cho, Nam Soo; Coudray, Clotilde; Dugoujon, Jean-Michel; Espinoza, Marta; González-Andrade, Fabricio; Hadi, Sibte; Immel, Uta-Dorothee; Marian, Catalin; Gonzalez-Martin, Antonio; Mertens, Gerhard (1 September 2011). "PopAffiliator: online calculator for individual affiliation to a major population group based on 17 autosomal short tandem repeat genotype profile". International Journal of Legal Medicine. 125 (5): 629–636. doi:10.1007/s00414-010-0472-2. hdl:10642/472. ISSN 1437-1596. PMID 20552217. S2CID 11740334.
- ↑ "Analysis of the short tandem repeat (STR) data published on Ramesses III and the Amarna ancient royal family (including Tutankhamun) showed a majority to have an affinity with “sub-Saharan” Africans in one affinity analysis, which does not mean that they lacked other affiliations—an important point that typological thinking obscures".Keita, S. O. Y. (September 2022). "Ideas about "Race" in Nile Valley Histories: A Consideration of "Racial" Paradigms in Recent Presentations on Nile Valley Africa, from "Black Pharaohs" to Mummy Genomest". Journal of Ancient Egyptian Interconnections. Retrieved 11 May 2023.
- 1 2 Drosou, Konstantina; et al. (2020). "The first reported case of the rare mitochondrial haplotype H4a1 in ancient Egypt". Scientific Reports. 10 (1): 17037. Bibcode:2020NatSR..1017037D. doi:10.1038/s41598-020-74114-9. PMC 7550590. PMID 33046824.
- 1 2 3 Bekada A, Fregel R, Cabrera VM, Larruga JM, Pestano J, Benhamamouch S, González AM (19 February 2013). "Introducing the Algerian mitochondrial DNA and Y-chromosome profiles into the North African landscape". PLOS ONE. 8 (2): e56775. Bibcode:2013PLoSO...856775B. doi:10.1371/journal.pone.0056775. PMC 3576335. PMID 23431392.
- 1 2 3 Yatsishina, E. B.; Bulygina, E. S.; Vasilyev, S. V.; Galeev, R. M.; Slobodova, N. V.; Tsygankova, S. V.; Sharko, F. S. (2020). "Paleogenetic Study of Ancient Mummies at the Kurchatov Institute". Nanotechnologies in Russia. 15 (9–10): 524–531. doi:10.1134/S1995078020050183. S2CID 232315321.
Haplogroup R1b1a1b isolated in the studied samples is also uncharacteristic of the modern Egyptian population; its frequency in the population is less than 1%, while it is found in approximately half of the male population of Western Europe.
- ↑ Cruciani F, La Fratta R, Trombetta B, Santolamazza P, Sellitto D, Colomb EB, Dugoujon JM, Crivellaro F, Benincasa T, Pascone R, Moral P, Watson E, Melegh B, Barbujani G, Fuselli S, Vona G, Zagradisnik B, Assum G, Brdicka R, Kozlov AI, Efremov GD, Coppa A, Novelletto A, Scozzari R (June 2007). "Tracing past human male movements in northern/eastern Africa and western Eurasia: new clues from Y-chromosomal haplogroups E-M78 and J-M12". Molecular Biology and Evolution. 24 (6): 1300–11. doi:10.1093/molbev/msm049. PMID 17351267.
- ↑ Battaglia, Vincenza; Fornarino, Simona; Al-Zahery, Nadia; Olivieri, Anna; Pala, Maria; Myres, Natalie M.; King, Roy J.; Rootsi, Siiri; Marjanovic, Damir; Primorac, Dragan; Hadziselimovic, Rifat; Vidovic, Stojko; Drobnic, Katia; Durmishi, Naser; Torroni, Antonio (2008). "Y-chromosomal evidence of the cultural diffusion of agriculture in Southeast Europe". European Journal of Human Genetics. 17 (6): 820–830. doi:10.1038/ejhg.2008.249. ISSN 1476-5438. PMC 2947100. PMID 19107149.
- 1 2 3 Manni F, Leonardi P, Barakat A, Rouba H, Heyer E, Klintschar M, McElreavey K, Quintana-Murci L (October 2002). "Y-chromosome analysis in Egypt suggests a genetic regional continuity in Northeastern Africa". Human Biology. 74 (5): 645–58. doi:10.1353/hub.2002.0054. PMID 12495079. S2CID 26741827.
- 1 2 3 4 Arredi B, Poloni ES, Paracchini S, Zerjal T, Fathallah DM, Makrelouf M, Pascali VL, Novelletto A, Tyler-Smith C (August 2004). "A predominantly neolithic origin for Y-chromosomal DNA variation in North Africa". American Journal of Human Genetics. 75 (2): 338–45. doi:10.1086/423147. PMC 1216069. PMID 15202071.
- ↑ Krings M, Salem AE, Bauer K, Geisert H, Malek AK, Chaix L, Simon C, Welsby D, Di Rienzo A, Utermann G, Sajantila A, Pääbo S, Stoneking M (April 1999). "mtDNA analysis of Nile River Valley populations: A genetic corridor or a barrier to migration?". American Journal of Human Genetics. 64 (4): 1166–76. doi:10.1086/302314. PMC 1377841. PMID 10090902.
- ↑ Underhill (2002), Bellwood and Renfrew, ed., Inference of Neolithic Population Histories using Y-chromosome Haplotypes, Cambridge: McDonald Institute for Archaeological Research, ISBN 978-1-902937-20-5.
- 1 2 Cruciani, Fulvio; La Fratta, Roberta; Trombetta, Beniamino; Santolamazza, Piero; Sellitto, Daniele; Colomb, Eliane Beraud; Dugoujon, Jean-Michel; Crivellaro, Federica; Benincasa, Tamara; Pascone, Roberto; Moral, Pedro; Watson, Elizabeth; Melegh, Bela; Barbujani, Guido; Fuselli, Silvia; Vona, Giuseppe; Zagradisnik, Boris; Assum, Guenter; Brdicka, Radim; Kozlov, Andrey I.; Efremov, Georgi D.; Coppa, Alfredo; Novelletto, Andrea; Scozzari, Rosaria (June 2007). "Tracing past human male movements in northern/eastern Africa and western Eurasia: new clues from Y-chromosomal haplogroups E-M78 and J-M12". Molecular Biology and Evolution. 24 (6): 1300–1311. doi:10.1093/molbev/msm049. ISSN 0737-4038. PMID 17351267.
- ↑ Luca Cavalli-Sforza L, Menozzi P, Piazza A (5 August 1996). The History and Geography of Human Genes. Princeton University Press. ISBN 978-0-691-02905-4.
- 1 2 Stevanovitch A, Gilles A, Bouzaid E, Kefi R, Paris F, Gayraud RP, Spadoni JL, El-Chenawi F, Béraud-Colomb E, et al. (January 2004). "Mitochondrial DNA sequence diversity in a sedentary population from Egypt". Annals of Human Genetics. 68 (Pt 1): 23–39. doi:10.1046/j.1529-8817.2003.00057.x. PMID 14748828. S2CID 44901197.
- ↑ Cavalli-Sforza, L.L., P. Menozzi, and A. Piazza (1994). The History and Geography of Human Genes Archived 4 April 2023 at the Wayback Machine. Princeton:Princeton University Press. p. 174. ISBN 0-691-08750-4
- ↑ Cavalli-Sforza, Luigi Luca; Cavalli-Sforza, Luca; Menozzi, Paolo; Piazza, Alberto (1994). The History and Geography of Human Genes. Princeton University Press. pp. 136 (Chapter 2). ISBN 978-0-691-08750-4. Archived from the original on 4 April 2023. Retrieved 19 March 2023.
- ↑ Kivisild T, Reidla M, Metspalu E, Rosa A, Brehm A, Pennarun E, Parik J, Geberhiwot T, Usanga E, Villems R (2004). "Ethiopian Mitochondrial DNA Heritage: Tracking Gene Flow Across and Around the Gate of Tears". American Journal of Human Genetics. 75 (5): 752–770. doi:10.1086/425161. PMC 1182106. PMID 15457403.
- ↑ Terreros, Maria C.; Martinez, Laisel; Herrera, Rene J. (2005). "Polymorphic Alu Insertions and Genetic Diversity Among African Populations". Human Biology. 77 (5): 675–704. doi:10.1353/hub.2006.0009. ISSN 0018-7143. JSTOR 41466364. PMID 16596946. S2CID 36880409.
- ↑ González AM, Larruga JM, Abu-Amero KK, Shi Y, Pestano J, Cabrera VM (July 2007). "Mitochondrial lineage M1 traces an early human backflow to Africa". BMC Genomics. 8: 223. doi:10.1186/1471-2164-8-223. PMC 1945034. PMID 17620140.
- ↑ Kivisild T, Rootsi S, Metspalu M, Mastana S, Kaldma K, Parik J, Metspalu E, Adojaan M, et al. (2003). "The Genetic Heritage of the Earliest Settlers Persists Both in Indian Tribal and Caste Populations". American Journal of Human Genetics. 72 (2): 313–32. doi:10.1086/346068. PMC 379225. PMID 12536373.
- ↑ Quintana-Murci L, Semino O, Bandelt HJ, Passarino G, McElreavey K, Santachiara-Benerecetti AS (December 1999). "Genetic evidence of an early exit of Homo sapiens sapiens from Africa through eastern Africa". Nature Genetics. 23 (4): 437–441. doi:10.1038/70550. ISSN 1061-4036. PMID 10581031. S2CID 2000627. Archived from the original on 19 April 2022. Retrieved 19 April 2022.
- 1 2 3 4 Keita SO (2005). "History in the interpretation of the pattern of p49a,f TaqI RFLP Y-chromosome variation in Egypt: a consideration of multiple lines of evidence". American Journal of Human Biology. 17 (5): 559–67. doi:10.1002/ajhb.20428. PMID 16136533. S2CID 33076762.
- ↑ Cruciani F, La Fratta R, Trombetta B, Santolamazza P, Sellitto D, Colomb EB, Dugoujon JM, Crivellaro F, Benincasa T, Pascone R, Moral P, Watson E, Melegh B, Barbujani G, Fuselli S, Vona G, Zagradisnik B, Assum G, Brdicka R, Kozlov AI, Efremov GD, Coppa A, Novelletto A, Scozzari R (June 2007). "Tracing past human male movements in northern/eastern Africa and western Eurasia: new clues from Y-chromosomal haplogroups E-M78 and J-M12". Molecular Biology and Evolution. 24 (6): 1300–11. doi:10.1093/molbev/msm049. PMID 17351267.
- ↑ Keita, S.O.Y. (ed Bengston, John) (3 December 2008). "Geography, selected Afro-Asiatic families, and Y Chromosome lineage variation: An exploration in linguistics and phylogeography" in In Hot Pursuit of Language in Prehistory: Essays in the four fields of anthropology. In honor of Harold Crane Fleming. John Benjamins Publishing. pp. 3–15. ISBN 978-90-272-8985-8.
{{cite book}}: CS1 maint: multiple names: authors list (link) - ↑ Babiker, Hiba MA; Schlebusch, Carina M.; Hassan, Hisham Y.; Jakobsson, Mattias (4 May 2011). "Genetic variation and population structure of Sudanese populations as indicated by 15 Identifiler sequence-tagged repeat (STR) loci". Investigative Genetics. 2 (1): 12. doi:10.1186/2041-2223-2-12. ISSN 2041-2223. PMC 3118356. PMID 21542921. S2CID 7390979.
- 1 2 Hollfelder, Nina; Schlebusch, Carina M.; Günther, Torsten; Babiker, Hiba; Hassan, Hisham Y.; Jakobsson, Mattias (24 August 2017). "Northeast African genomic variation shaped by the continuity of indigenous groups and Eurasian migrations". PLOS Genetics. 13 (8): e1006976. doi:10.1371/journal.pgen.1006976. PMC 5587336. PMID 28837655.
- 1 2 Taha, Tarek; Elzalabany, Sagy; Fawzi, Sahar; Hisham, Ahmed; Amer, Khaled; Shaker, Olfat (1 August 2020). "Allele frequency comparative study between the two main Egyptian ethnic groups". Forensic Science International. 313: 110348. doi:10.1016/j.forsciint.2020.110348. ISSN 0379-0738. PMID 32521421. S2CID 219586129. Archived from the original on 3 June 2021. Retrieved 14 April 2023.
- ↑ Lucotte G, Mercier G (May 2003). "Brief communication: Y-chromosome haplotypes in Egypt". American Journal of Physical Anthropology. 121 (1): 63–6. doi:10.1002/ajpa.10190. PMID 12687584.
- ↑ Keita, S.O.Y. (September 2005). "History in the interpretation of the pattern of p49a,fTaqI RFLP Y-chromosome variation in Egypt: A consideration of multiple lines of evidence". American Journal of Human Biology. 17 (5): 559–567. doi:10.1002/ajhb.20428. ISSN 1042-0533. PMID 16136533. S2CID 33076762. Archived from the original on 11 June 2022. Retrieved 11 June 2022.
- ↑ Dugoujon J.M., Coudray C., Torroni A., Cruciani F., Scozzari F., Moral P., Louali N., Kossmann M. The Berber and the Berbers: Genetic and linguistic diversities
- ↑ Saunier JL, Irwin JA, Strouss KM, Ragab H, Sturk KA, Parsons TJ (June 2009). "Mitochondrial control region sequences from an Egyptian population sample". Forensic Science International: Genetics. 3 (3): E97–E103. doi:10.1016/j.fsigen.2008.09.004. PMID 19414160.
- ↑ Mohammad, T.; Xue, Yali; Evison, M.; Tyler-Smith, Chris (November 2009). "Genetic structure of nomadic Bedouin from Kuwait". Heredity. 103 (5): 425–433. doi:10.1038/hdy.2009.72. ISSN 0018-067X. PMC 2869035. PMID 19639002.
- ↑ Serra-Vidal, Gerard; Lucas-Sanchez, Marcel; Fadhlaoui-Zid, Karima; Bekada, Asmahan; Zalloua, Pierre; Comas, David (18 November 2019). "Heterogeneity in Palaeolithic Population Continuity and Neolithic Expansion in North Africa". Current Biology. 29 (22): 3953–3959.e4. doi:10.1016/j.cub.2019.09.050. PMID 31679935. S2CID 204972040.
- ↑ Crubézy, Eric (January 2010). "Le peuplement de la vallée du Nil". Archeo-Nil (in French). 20 (1): 25–42. doi:10.3406/arnil.2010.999. S2CID 248278173.
- ↑ Crubézy, Eric (January 2010). "Le peuplement de la vallée du Nil". Archeo-Nil (in French). 20 (1): 25–42. doi:10.3406/arnil.2010.999. S2CID 248278173.
- 1 2 Hassan HY, Underhill PA, Cavalli-Sforza LL, Ibrahim ME (November 2008). "Y-chromosome variation among Sudanese: restricted gene flow, concordance with language, geography, and history". American Journal of Physical Anthropology. 137 (3): 316–23. doi:10.1002/ajpa.20876. PMID 18618658.
- ↑ Trombetta B, D'Atanasio E, Massaia A, Ippoliti M, Coppa A, Candilio F, Coia V, Russo G, Dugoujon JM, Moral P, Akar N, Sellitto D, Valesini G, Novelletto A, Scozzari R, Cruciani F (June 2015). "Phylogeographic Refinement and Large Scale Genotyping of Human Y Chromosome Haplogroup E Provide New Insights into the Dispersal of Early Pastoralists in the African Continent". Genome Biology and Evolution. 7 (7): 1940–50. doi:10.1093/gbe/evv118. PMC 4524485. PMID 26108492.
- ↑ Yousif, Hisham; Eltayeb, Muntaser (July 2009). Genetic Patterns of Y-chromosome and Mitochondrial DNA Variation, with Implications to the Peopling of the Sudan (Thesis). Archived from the original on 12 November 2021. Retrieved 12 November 2021.
- 1 2 Dobon B, Hassan HY, Laayouni H, Luisi P, Ricaño-Ponce I, Zhernakova A, Wijmenga C, Tahir H, Comas D, Netea MG, Bertranpetit J (May 2015). "The genetics of East African populations: a Nilo-Saharan component in the African genetic landscape". Scientific Reports. 5: 9996. Bibcode:2015NatSR...5E9996D. doi:10.1038/srep09996. PMC 4446898. PMID 26017457.
| Sub-topics | |
|---|---|
| Genetic history by region | |
| Population genetics by group |
|
| |