
Dwarf elephants are prehistoric members of the order Proboscidea which, through the process of allopatric speciation on islands, evolved much smaller body sizes (around 1–2.3 metres (3 ft 3 in – 7 ft 7 in)) in comparison with their immediate ancestors. Dwarf elephants are an example of insular dwarfism, the phenomenon whereby large terrestrial vertebrates (usually mammals) that colonize islands evolve dwarf forms, a phenomenon attributed to adaptation to resource-poor environments and lack of predation and competition. Some modern populations of Asian elephants have also undergone size reduction on islands to a lesser degree, resulting in populations of pygmy elephants.
Fossil remains of dwarf elephants have been found on the Mediterranean islands of Cyprus, Malta (at Għar Dalam), Crete (in Chania at Vamos, Stylos and in a now-underwater cave on the coast), Sicily, Sardinia, the Cyclades Islands and the Dodecanese Islands, which are mostly members of the genus Palaeoloxodon, descending from the large 4 metres (13 ft) tall mainland European species Palaeoloxodon antiquus. Dwarf species of elephants and Stegodon have been found on the islands of Indonesia and the Philippines. The Channel Islands of California once supported a dwarf species descended from Columbian mammoths,[1] while the woolly mammoths that existed on Wrangel Island north of Siberia were once considered dwarfs, but are not anymore.
The Mediterranean islands
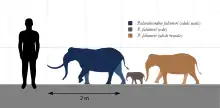
Dwarf elephants first inhabited the Mediterranean islands during the Pleistocene, including all the major islands with the apparent exception of Corsica and the Balearics. Mediterranean dwarf elephants have generally been considered as members of the genus Palaeoloxodon, derived from the continental straight-tusked elephant, Palaeoloxodon antiquus (Falconer & Cautley, 1847), Syn.: Elephas antiquus. An exception is the dwarf Middle-Late Pleistocene Sardinian mammoth, Mammuthus lamarmorai (Major, 1883), the first endemic elephant of the Mediterranean islands recognized as belonging to the mammoth line. Mammuthus creticus from the Early Pleistocene of Crete, formerly considered a member of Palaeoloxodon, is now also considered to be a mammoth, and approaches the size of the smallest dwarf elephants.[3]
During low sea levels, the Mediterranean islands were colonised again and again, giving rise, sometimes on the same island, to several species (or subspecies) of different body sizes. As the Ice Age came to an end, sea levels rose, stranding elephants on the island. The island of Sicily appears to have been colonised by proboscideans in at least three separate waves of colonisation. These endemic dwarf elephants were taxonomically different on each island or group of very close islands, like the Cyclades archipelago.
There are many uncertainties about the time of colonisation, the phylogenetic relationships and the taxonomic status of dwarf elephants on the Mediterranean islands. Extinction of the insular dwarf elephants has not been correlated with the arrival of humans to the islands. Furthermore, it has been suggested by the palaeontologist Othenio Abel in 1914,[4] that the finding of skeletons of such elephants sparked the idea that they belonged to giant one-eyed monsters, because the center nasal opening was thought to be the socket of a single eye, and thus perhaps were, for example, the origin of the one-eyed Cyclopes of Greek mythology.
Italy and Malta

| Island | Taxon | Author |
|---|---|---|
| Sardinia | Mammuthus lamarmorai | (Major, 1883) |
| Malta | Palaeoloxodon falconeri | (Busk, 1869) |
| Palaeoloxodon mnaidriensis | (Adams, 1874) | |
| Sicily | Palaeoloxodon falconeri | (Busk, 1869) |
| Palaeoloxodon cf. mnaidriensis | (Adams, 1874) |
Greece
Crete

| Island | Taxon | Author |
|---|---|---|
| Crete | Mammuthus creticus | (Bate, 1907) |
| Palaeoloxodon creutzburgi | (Kuss, 1965) |
Poulakakis and others proposed in 2002 to rename all the described specimens of larger size than Mammuthus creticus under the new subspecies name Elephas antiquus creutzburgi (Kuss, 1965).[6] After DNA research, published in 2006, it has been proposed to rename Elephas (Palaeoloxodon) creticus into Mammuthus creticus (Bate, 1907). In a recent study of 2007, it was argued for the groundlessness of the theory by Poulakakis et al. in 2006,[7] showing the weak points of that DNA research.[8] However, morphological data is at least equivocal, and may also support placement in Mammuthus.
Mammuthus creticus is the earlier species known from the Early Pleistocene. It is the smallest mammoth[9] and is among the smallest dwarf elephants known, with a shoulder height of about 1 metre (3.3 ft) and a weight of about 180 kilograms (400 lb).[10] Palaeoloxodon creutzburgi from the Middle Pleistocene and Late Pleistocene is significantly larger, with an estimated body mass comparable to living Asian elephant, around 40% the size of its mainland ancestor.[11]
Cyclades
| Island | Taxon | Author |
|---|---|---|
| Delos | Palaeoloxodon sp. | Vaufrey, 1929 |
| Naxos | Palaeoloxodon lomolinoi | Van der Geer et al., 2014[12] |
| Paros | Elephantidae indet. | Georgalas, 1929 |
| Kythnos | Elephantidae indet. | Honea, 1975 |
| Milos | Elephantidae indet. | Papp, 1953 |
| Serifos | Elephantidae indet. | Papp, 1953 |
Remains of paleoloxodontine elephants have been reported from several Cyclades islands. The Delos elephant's size is comparable to Palaeoloxodon mnaidriensis.[12] The Naxos elephant has been described as Palaeoloxodon lomolinoi and it was smaller than the Delos elephant with a calculated body mass of only 10% of that of its mainland ancestor Palaeoloxodon antiquus.[12]
Dodecanese
| Island | Taxon | Author |
|---|---|---|
| Astypalaia | Palaeoloxodon sp. | Athanassiou et al., 2019[2] |
| Kasos | Palaeoloxodon aff. creutzburgi | Sen et al., 2014[13] |
| Rhodes | Palaeoloxodon sp. | Symeonides et al., 1974 |
| Tilos | Palaeoloxodon tiliensis | (Theodorou et al. 2007)[14] |
On the island of Rhodes, bones of an endemic dwarf elephant have been discovered. This elephant was similar in size to Palaeoloxodon mnaidriensis.
Two groups of remains of dwarf elephants have been found on the island of Tilos. They are similar in size to Palaeoloxodon mnaidriensis and the smaller Palaeoloxodon falconeri, but the two groups indicate sexual dimorphism.[15][16] The remains had originally been designated to Palaeoloxodon antiquus falconeri (Busk, 1867). However, this name refers to the dwarf elephants from the island of Malta. As a result, since no migration route between the two islands can be proven, this name should not be used when referring to the elephant remnants from Tilos. The species has been described as Elephas tiliensis in 2007[14] and it is now assigned to genus Palaeoloxodon.
The Tilos dwarf elephant is the first dwarf elephant whose DNA sequence has been studied. The results of this research are consistent with previous morphological reports, according to which Palaeoloxodon is more closely related to Elephas than to Loxodonta or Mammuthus.[17] On the other hand, DNA analysis in 2017 conclude that the closest living relative of the genus Palaeoloxodon is the African forest elephant (Loxodonta cyclotis).[18] The paper suggests that the current view of elephant evolution is "in need of substantial revision".[18] After the study of new osteological material[14] that has been excavated in anatomical connection in the Charkadio Cave on Tilos island the new species name Palaeoloxodon tiliensis has been assigned to the Tilos dwarf elephants.
The Tilos dwarf elephant is the only among the Mediterranean elephants that has been proven to have survived into the Holocene, with the most recent remains having been dated to just 3040-1840 BCE.[19]
Cyprus
| Island | Taxon | Author |
|---|---|---|
| Cyprus | Palaeoloxodon cypriotes | (Bate, 1903)[20] |
| Palaeoloxodon xylophagou | Athanassiou et al., 2015[21] |
The Cyprus dwarf elephant survived at least until 11,000 BC. Its estimated body weight was only 200 kg (440 lb), only 2% of its 10,000 kg (22,000 lb) ancestor. Molars of this dwarf elephant are reduced to approximately 40% the size of mainland straight-tusked elephants.
Remains of the species were first discovered and recorded by Dorothea Bate in a cave in the Kyrenia hills of Cyprus in 1902 and reported in 1903.[20][22]
The Channel Islands of California
The Columbian mammoth (Mammuthus columbi) produced a separate, isolated population at the end of the Pleistocene on the Channel Islands of California, most likely about 40,000 years ago (although the time of isolation is not fully known). Selective forces on the Channel Islands resulted in smaller animals, forming a new species, the pygmy mammoth (Mammuthus exilis).[23] Channel Islands mammoths ranged from 150–190 cm (59–75 in) in shoulder height.
Wrangel Island
During the last ice age, woolly mammoths (Mammuthus primigenius) lived on Wrangel Island in the Arctic Ocean and survived until 1700 BC, the most recent survival of any known mammoth population. Wrangel Island is thought to have become separated from the mainland by 12000 BC. It was assumed that Wrangel Island mammoths ranged from 180–230 cm (71–91 in) in shoulder height and were for a time considered "dwarf mammoths".[24] However this classification has been re-evaluated and since the Second International Mammoth Conference in 1999, these mammoths are no longer considered to be true "dwarf mammoths".[25]
Indonesia and the Philippines
In Indonesia and the Philippines, evidence of a succession of distinct endemic island faunas has been found, including dwarfed elephants and species of Stegodon.
Flores

During the late Early Pleistocene, Flores was inhabited by the dwarf species Stegodon sondaarii, around 15% of the size of mainland Stegodon species, which was around 120 cm (3.9 ft) tall at the shoulder and weighed about 350–400 kilograms (770–880 lb). This species became extinct around 1 million years ago,[10][26][27] being replaced by Stegodon florensis. Stegodon florensis shows a progressive size reduction with time, with the earlier Middle Pleistocene subspecies Stegodon florensis florensis estimated to be around 50% the size of mainland Stegodon species with a shoulder height of around 190 cm (6.2 ft) and a body mass of around 1.7 tons, while the later Stegodon florensis insularis from the Late Pleistocene is estimated to be around 17% the size of mainland Stegodon species, with a shoulder height of around 130 cm (4.3 ft), and a body mass of about 570 kilograms (1,260 lb) [27][28] Stegodon florensis became extinct about 50,000 years ago, around the time of the arrival of modern humans to Flores.[26]
Sulawesi
During the Late Pliocene-Early Pleistocene on Sulawesi, two species of dwarf proboscidean coinhabited the island, the elephant Stegoloxodon celebensis, and Stegodon sompoensis.[29] The former was about 150 cm (4.9 ft) tall,[30] while the latter was around 32% the size of mainland Stegodon species, with an estimated body mass of about a ton.[27] Later in the Pleistocene, these animals were replaced by larger-sized species of Stegodon and elephants,[29][31] with an indeterminate Stegodon species from the Middle Pleistocene of Sulawesi being around 57% the size of mainland species, with an estimated bodymass of about 2 tons.[27]
Java
The species Stegodon trigonocephalus is known from the Early-Middle Pleistocene of Java.[32] A population from the Trinil H.K locality, which likely dates to the Middle Pleistocene,[33] is around 65% the size of mainland Stegodon species.[27] Large individuals are estimated to have reached around 280 cm (9.2 ft) at the shoulders, with a body mass of around 5 tons.[10] Other smaller unnamed Stegodon species are also known from the Early Pleistocene on the island.[32] The extinct dwarf elephant species Stegoloxodon indonesicus is also known from the Early Pleistocene of Java, which is probably closely related to S. celebensis from Sulawesi, but whose relationships to other elephants are obscure.[34][32]
Sumba
The species Stegodon sumbaensis of an uncertain Middle-Late Pleistocene age from Sumba is one the smallest known species, at around 8% of the size of its mainland ancestor, with an estimated body mass of around 250 kilograms (550 lb).[27]
Timor
The species S. timorensis is known from the Middle Pleistocene of Timor. It is a small-sized species, only slightly larger than S. sondaarii,[35] and around 23% the size of mainland species, with an estimated body mass of around 770 kilograms (1,700 lb).[27]
Luzon
On Luzon the dwarf Stegodon luzonensis is known from remains found in the Manila Basin of an uncertain Pleistocene age,[36] as well as remains found near the early Middle Pleistocene Nesorhinus butchery site.[37] It is around 40% the size of mainland Stegodon species, with a body mass of around 1.3 tons.[27] The extinct dwarf elephant Elephas beyeri is also known from the island,[27] which is estimated to have been about 1.2 metres (3.9 ft) in shoulder height.[38]
Mindanao
On the island of Mindanao, the dwarf Stegodon species Stegodon mindanensis was present at some point in the Pleistocene. It has an estimated body mass of around 400 kilograms (880 lb).[39]
See also
References
- ↑ "The Pygmy Mammoth (U.S. National Park Service)".
- 1 2 Athanassiou, A.; Van der Geer, A. A. E.; Lyras, G. A. (2019). "Pleistocene insular Proboscidea of the Eastern Mediterranean: A review and update". Quaternary Science Reviews. 218: 306–321. doi:10.1016/j.quascirev.2019.06.028. S2CID 199107354.
- ↑ Herridge, Victoria L.; Lister, Adrian M. (2012-05-09). "Extreme insular dwarfism evolved in a mammoth". Proceedings of the Royal Society B: Biological Sciences. 279 (1741): 3193–3200. doi:10.1098/rspb.2012.0671. ISSN 0962-8452. PMC 3385739. PMID 22572206.
- ↑ Abel's surmise is noted by Adrienne Mayor in The First Fossil Hunters: Paleontology in Greek and Roman Times (Princeton University Press) 2000. [See illus. ed., 2001: ISBN 0691089779]
- ↑ Palombo, M. R.; Antonioli, F.; Di Patti, C.; Presti, V. L.; Scarborough, M. E. (2020). "Was the dwarfed Palaeoloxodon from Favignana Island the last endemic Pleistocene elephant from the western Mediterranean islands?". Historical Biology. 33 (10): 1–19. doi:10.1080/08912963.2020.1772251. S2CID 225710152.
- ↑ Poulakakis, N.; Mylonas, M.; Lymberakis, P.; and Fassoulas, C. (2002-10-01). "Origin and taxonomy of the fossil elephants of the island of Crete (Greece): problems and perspectives". Palaeogeography, Palaeoclimatology, Palaeoecology. 186 (1–2): 163–183. doi:10.1016/S0031-0182(02)00451-0.
- ↑ Poulakakis N.; Parmakelis A.; Lymberakis P.; Mylonas M.; Zouros E.; Reese D.; Glaberman S.; Caccone A. (2006). "Ancient DNA forces reconsideration of evolutionary history of Mediterranean pygmy elephantids". Biol. Lett. 2 (3): 451–454. doi:10.1098/rsbl.2006.0467. PMC 1686204. PMID 17148428.
- ↑ Orlando, L.; Pagés, M.; Calvignac, S.; et al. (2007-02-22). "Does the 43bp sequence from an 800000 year old Cretan dwarf elephantid really rewrite the textbook on mammoths?". Biology Letters. 3 (1): 57–59. doi:10.1098/rsbl.2006.0536. PMC 2373798. PMID 17443966.
- ↑ Herridge, V. L.; Lister, A. M. (2012). "Extreme insular dwarfism evolved in a mammoth". Proceedings of the Royal Society B: Biological Sciences. 279 (1741): 3193–3300. doi:10.1098/rspb.2012.0671. PMC 3385739. PMID 22572206.
- 1 2 3 Larramendi, A. (2016). "Shoulder height, body mass and shape of proboscideans" (PDF). Acta Palaeontologica Polonica. 61. doi:10.4202/app.00136.2014. S2CID 2092950.
- ↑ Athanassiou, Athanassios; van der Geer, Alexandra A.E.; Lyras, George A. (August 2019). "Pleistocene insular Proboscidea of the Eastern Mediterranean: A review and update". Quaternary Science Reviews. 218: 306–321. Bibcode:2019QSRv..218..306A. doi:10.1016/j.quascirev.2019.06.028. S2CID 199107354.
- 1 2 3 4 Van der Geer, A. A. E.; Lyras, G. A.; Van den Hoek Ostende, L. W.; De Vos, J.; Drinia, H. (2014). "A dwarf elephant and a rock mouse on Naxos (Cyclades, Greece) with a revision of the palaeozoogeography of the Cycladic Islands (Greece) during the Pleistocene". Palaeogeography, Palaeoclimatology, Palaeoecology. 404: 133–144. doi:10.1016/j.palaeo.2014.04.003. hdl:10795/3263.
- ↑ Sen, S.; Barrier, E.; Crété, X. (2014). "Late Pleistocene Dwarf Elephants from the Aegean Islands of Kassos and Dilos, Greece". Annales Zoologici Fennici. 51 (1–2): 27–42. doi:10.5735/086.051.0204. S2CID 85190966.
- 1 2 3 Theodorou, G.E.; Symeonides, N.; Stathopoulou, E. (2007). "Elephas tiliensis n. sp. from Tilos island (Dodecanese, Greece)". Hellenic Journal of Geosciences. 42: 19–32.
- ↑ Theodorou, G. (1983). The dwarf elephants of the Charkadio cave on the island of Tilos (Dodekanese, Greece). Phd Thesis Athens University. p. 321 pp.
- ↑ Theodorou, G.E. (1988). "Environmental factors affecting the evolution of islands endemics: The Tilos example for Greece". Modern Geology. 13: 183–188.
- ↑ Poulakakis, Nikos; Theodorou, Georgios E.; Zouros, Eleftherios; Mylonas, Moysis (2002-09-01). "Molecular Phylogeny of the Extinct Pleistocene Dwarf Elephant Palaeoloxodon antiquus falconeri from Tilos Island, Dodekanisa, Greece". Journal of Molecular Evolution. Springer Science and Business Media LLC. 55 (3): 364–374. doi:10.1007/s00239-002-2337-x. ISSN 0022-2844. PMID 12187389. S2CID 21140948.
- 1 2 Meyer, Matthias; Palkopoulou, Eleftheria; Baleka, Sina; Stiller, Mathias; Penkman, Kirsty E H; Alt, Kurt W; Ishida, Yasuko; Mania, Dietrich; Mallick, Swapan; Meijer, Tom; Meller, Harald; Nagel, Sarah; Nickel, Birgit; Ostritz, Sven; Rohland, Nadin; Schauer, Karol; Schüler, Tim; Roca, Alfred L; Reich, David; Shapiro, Beth; Hofreiter, Michael (6 June 2017). "Palaeogenomes of Eurasian straight-tusked elephants challenge the current view of elephant evolution". eLife. 6: e25413. doi:10.7554/eLife.25413. PMC 5461109. PMID 28585920.
- ↑ Masseti, M. (2008). The most ancient explorations of the Mediterranean. Proc. Calif. Acad. Sci. 4th Ser, 59(Suppl I), 1-18.
- 1 2 Bate, D. M. A.: Preliminary Note on the Discovery of a Pigmy Elephant in the Pleistocene of Cyprus in Proceedings of the Royal Society of London Vol. 71 (1902 - 1903), pp. 498-500
- ↑ Athanassiou, A.; Herridge, V.; Reese, D. S.; Iliopoulos, G.; Roussiakis, S.; Mitsopoulou, V.; Tsiolakis, E.; Theodorou, G. (2015). "Cranial evidence for the presence of a second endemic elephant species on Cyprus". Quaternary International. 379: 47–57. doi:10.1016/j.quaint.2015.05.065.
- ↑ Dorothea Bate, Cyprus work diary 1901–02, 3 volumes, Natural History Museum's earth sciences library, palaeontology MSS
- ↑ Rocha, Veronica (16 September 2016). "Well-preserved mammoth skull unearthed on Channel Islands puzzles scientists". Los Angeles Times. Retrieved 24 September 2016.
- ↑ Vartanyan, S.L.; Garutt, V.E.; Sher, A.V. (1993). "Holocene dwarf mammoths from Wrangel Island in the Siberian Arctic". Nature. 362 (6418): 337–340. doi:10.1038/362337a0. PMID 29633990. S2CID 4249191.
- ↑ Tikhonov, Alexei; Larry Agenbroad; Sergey Vartanyan (2003). "Comparative analysis of the mammoth populations on Wrangel Island and the Channel Islands". Deinsea. 9: 415–420. ISSN 0923-9308. Archived from the original on 2012-06-11.
- 1 2 van den Bergh, Gerrit D.; Alloway, Brent V.; Storey, Michael; Setiawan, Ruly; Yurnaldi, Dida; Kurniawan, Iwan; Moore, Mark W.; Jatmiko; Brumm, Adam; Flude, Stephanie; Sutikna, Thomas; Setiyabudi, Erick; Prasetyo, Unggul W.; Puspaningrum, Mika R.; Yoga, Ifan (October 2022). "An integrative geochronological framework for the Pleistocene So'a basin (Flores, Indonesia), and its implications for faunal turnover and hominin arrival". Quaternary Science Reviews. 294: 107721. doi:10.1016/j.quascirev.2022.107721. hdl:10072/418777. S2CID 252290750.
- 1 2 3 4 5 6 7 8 9 Geer, Alexandra A. E.; Bergh, Gerrit D.; Lyras, George A.; Prasetyo, Unggul W.; Due, Rokus Awe; Setiyabudi, Erick; Drinia, Hara (August 2016). "The effect of area and isolation on insular dwarf proboscideans". Journal of Biogeography. 43 (8): 1656–1666. doi:10.1111/jbi.12743. ISSN 0305-0270. S2CID 87958022.
- ↑ Puspaningrum, Mika; Van Den Bergh, Gerrit; Chivas, Allan; Setiabudi, Erick; Kurniawan, Iwan; Brumm, Adam; and Sutikna, Thomas, "Preliminary results of dietary and environmental reconstructions of Early to Middle Pleistocene Stegodons from the So'a Basin of Flores, Indonesia, based on enamel stable isotope records" (2014). Faculty of Science, Medicine and Health - Papers: part A. 2035.
- 1 2 Vos, John de; Ostende, Lars W. van den Hoek; Bergh, Gert D. van den (2007), Renema, Willem (ed.), "Patterns in Insular Evolution of Mammals: A Key to Island Palaeogeography", Biogeography, Time, and Place: Distributions, Barriers, and Islands, Topics in Geobiology, Dordrecht: Springer Netherlands, vol. 29, pp. 315–345, doi:10.1007/978-1-4020-6374-9_10, ISBN 978-1-4020-6373-2, retrieved 2023-08-10
- ↑ Alink, Gerrit; Adhityatama, Shinatria; Simanjuntak, Truman (2017-12-29). "The Descriptive Analysis of Palaeolithic Stone Tools from Sulawesi, Collected by the Indonesian-Dutch Expedition in 1970". AMERTA. 35 (2): 75. doi:10.24832/amt.v35i2.252.
- ↑ Brumm, Adam; Hakim, Budianto; Ramli, Muhammad; Aubert, Maxime; van den Bergh, Gerrit D.; Li, Bo; Burhan, Basran; Saiful, Andi Muhammad; Siagian, Linda; Sardi, Ratno; Jusdi, Andi; Abdullah; Mubarak, Andi Pampang; Moore, Mark W.; Roberts, Richard G. (2018-04-11). Hardy, Karen (ed.). "A reassessment of the early archaeological record at Leang Burung 2, a Late Pleistocene rock-shelter site on the Indonesian island of Sulawesi". PLOS ONE. 13 (4): e0193025. doi:10.1371/journal.pone.0193025. ISSN 1932-6203. PMC 5894965. PMID 29641524.
- 1 2 3 Puspaningrum, Mika R.; van den Bergh, Gerrit D.; Chivas, Allan R.; Setiabudi, Erick; Kurniawan, Iwan (January 2020). "Isotopic reconstruction of Proboscidean habitats and diets on Java since the Early Pleistocene: Implications for adaptation and extinction". Quaternary Science Reviews. 228: 106007. doi:10.1016/j.quascirev.2019.106007. S2CID 212876762.
- ↑ L. Hilgen, Sander; Pop, Eduard; Adhityatama, Shinatria; A. Veldkamp, Tom; W.K. Berghuis, Harold; Sutisna, Indra; Yurnaldi, Dida; Dupont-Nivet, Guillaume; Reimann, Tony; Nowaczyk, Norbert; F. Kuiper, Klaudia; Krijgsman, Wout; B. Vonhof, Hubert; Ekowati, Dian Rahayu; Alink, Gerrit (February 2023). "Revised age and stratigraphy of the classic Homo erectus-bearing succession at Trinil (Java, Indonesia)". Quaternary Science Reviews. 301: 107908. doi:10.1016/j.quascirev.2022.107908.
- ↑ Markov, Georgi N.; Saegusa, Haruo (2008-09-01). "On the validity of Stegoloxodon Kretzoi, 1950 (Mammalia: Proboscidea)". Zootaxa. 1861 (1): 55. doi:10.11646/zootaxa.1861.1.5. ISSN 1175-5334.
- ↑ Louys, Julien; Price, Gilbert J.; O’Connor, Sue (2016-03-10). "Direct dating of Pleistocene stegodon from Timor Island, East Nusa Tenggara". PeerJ. 4: e1788. doi:10.7717/peerj.1788. ISSN 2167-8359. PMC 4793331. PMID 26989625.
- ↑ von Koenigswald GHR. 1956. Fossil mammals from the Philippines. Quezon City: National Research Council of the Philippines, 1–14
- ↑ Ingicco, T.; van den Bergh, G. D.; Jago-on, C.; Bahain, J.-J.; Chacón, M. G.; Amano, N.; Forestier, H.; King, C.; Manalo, K.; Nomade, S.; Pereira, A.; Reyes, M. C.; Sémah, A.-M.; Shao, Q.; Voinchet, P. (May 2018). "Earliest known hominin activity in the Philippines by 709 thousand years ago". Nature. 557 (7704): 233–237. doi:10.1038/s41586-018-0072-8. ISSN 0028-0836. PMID 29720661. S2CID 13742336.
- ↑ Koenigswald, G.H.R. (1956). "Fossil mammals from the Philippines". Proceedings of the Fourth Far-Eastern Prehistory Congress.
- ↑ Ong, Perry (1998). "The Philippine Menagerie". The Philippine Archipelago. Makati city, Philippines: Asia Publishing Co. Ltd. pp. 227–255.