| Budoš Limestone | |
|---|---|
| Stratigraphic range: Lower Toarcian ~ | |
 Duga Valley, where the southernmost Budoš Limestone emerges | |
| Type | Geological formation |
| Thickness | 50–60 m (160–200 ft)[1] |
| Lithology | |
| Primary | Limestones alternated with green marls and calcareous stone |
| Other | Lithified limestone |
| Location | |
| Coordinates | 42°59′30″N 18°54′20″E / 42.99167°N 18.90556°E |
| Region | Nikšić |
| Country | |
| Type section | |
| Named for | Budoš Mountain |
| Named by | Pantić |
| Year defined | 1952[1] |
 Budoš Limestone (Montenegro) | |
The Budoš Limestone ("Budos Mountain Limestone", also known as Briska Breccia[2]) is a geological formation in Montenegro, dating to 180 million years ago, and covering the Toarcian stage of the Jurassic Period. It has been considered an important setting in Balkan paleontology, as it represents a unique terrestrial setting with abundant plant material, one of the few know from the Toarcian of Europe.[3] It is the regional equivalent to the Toarcian units of Spain such as the Turmiel Formation, units like the Azilal Formation of Morocco and others from the Mediterranean such as the Posidonia Beds of Greece and the Marne di Monte Serrone of Italy.[4] In the Adriatic section, this unit is an equivalent of the Calcare di Sogno of north Italy, as well represents almost the same type of ecosystem recovered in the older (Pliensbachian) Rotzo Formation of the Venetian region, know also for its rich floral record.[5]
Description
The Toarcian paleogeography of Montenegro was characterised by two major units, mostly found in the Dinarides: the High Karst Zone, representing a Carbonate Platform, concretely the Adriatic-Dinaric Carbonate Platform and the Budva Basin, that represented a pelagic setting where ammonites are abundant. The previous Pliensbachian Platform suffered in the Toarcian a partial flooding, with the carbonate facies recovered at W of Nikšić, NE of Podgorica and in the Rumija Mt remaining as environments close to the marginal part.[4] The Budva basin evolution in the Toarcian was marked by the changes in the sea level, developing a distally steepened ramp until the Lower Toarcian, and an accretionary rimmed platform in younger layers.[6] The Adriatic-Dinaric Carbonate Platform is well measured at the Mount Rumija where the transitional facies between the platform setting and the deeper pelagic environment is seen, recovering a lateral transition from a lagoonal environment exposed in Seoce to the platform edge, exposed in Tejani (called Tejani section), and finally the deeper water environment, called Livari section can be observed at the own Mount Rumija.[7]
The Seoce Section is likely linked with the Budoš Limestone depositional setting, found mostly on the mountain of the same name on the Dinarides near Nikšić. The main unit is lithologically almost identical to the major fossiliferous levels of the Rotzo Formation, composed by bituminous limestones and marly limestones (fenestrate limestones and tempestites) with several episodes of emersions, all of coastal origin and rich in plant detritus and leaf remains, connected to the typical Lithiotis reefs found in the Pliensbachian-Toarcian carbonate platforms in the Adriatic region.[5] The Budoš Limestone was delimited as younger than the Rotzo Formation due to its floral composition and the fact is overlain by the Late Toarcian-Aalenian greenish local claystone-limestone layers.[5]
The unit is mostly known by its rich macroflora, the most complete and only known of the Mediterranean Toarcian realm, with several characteristics, such as the abundant presence of thermophilic Bennettitales and the dominance of the Seed Fern Pachypteris, that grew on semi-arid climates.[8] Most of the research of the flora was done by Pantic between 1952 & 1981, recovering abundant Macroflora and Palynomorphs. Several genera where recovered, such as Coniopteris (Dicksoniaceae), Lindleycladus (Krassiloviaceae) and Elatides (Taxodiaceae).[9]
Ecosystem

It was considered initially that this flora grew in a continental setting, appearing on deposits that resemble modern inland deposition on ferric soils, thus, in a large inland valley with semi-arid conditions but with nearby large water bodies such as lakes.[10] Latter however, was interpreted that this flora developed on an island setting in the Dinaric Carbonate Platform, likely linked with the exposed layers of Seoce. This setting would be made of the emerged Budoshi High, representing an island flora; a humid belt would have existed along the shore, while coniferous vegetation would have prevailed in the drier interior.[11] The Budoš flora, as well Rumija and Seoce lithiotis facies were made after the Livari Supersequence created a massive lagoon in the inner ramp.[6] A common facies in the 3 locations shows about 1–2 m thick lagoon parasequences, from lithiotis rich subtidal packstone to shallower wackestone, where the lagoonal shale facies recovering the flora deposited.[6]
The main consensus is that the layers rich in flora belong to a Bahamian-type Mangrove system developed on a coastal setting with a nearby arid inland setting dominated by Cheirolepidiaceae conifers and Bennettites, that was either a submarine intraoceanic carbonate platform or part of a large landmass. The Mangrove system was mostly composed of seed ferns bearing the leaf genus Pachypteris linked with complex root systems that cover most of the layers, developed over and linked with the local aberrant bivalve (Lithiotis) reefs and evolved as a belt around the coast, yet is unknown how far reached.[10] The inland setting was dry and with common wildfire activity, as proven by the great amount of charcoal recovered in some of the layers.[10] The Lithiotis layers are intercalated by oolitic and oncolitic layers of likely subtidal origin, with several coastal cycles measured, such as development of lagoons and complete flooding of the vegetation levels, as well small coal-dominated sections. The ingression-regression trend allowed the development of the local mangroves.[10]
The same type of ecosystem was also recovered more recently on slightly older (Late Pliensbachian) rocks on Albania, with also great dominance of the genus Pachypteris linked with root systems along Lithiotis reefs, with evidence of catastrophic events which “killed” the flora.[12] These types of layers have been vinculated with the early evolution of crabs.[13]
Fossil content
Flora
Palynology
Color key
|
Notes Uncertain or tentative taxa are in small text; |
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
Nannoceratopsis[9] |
|
|
Cysts |
A Dinoflajellate, member of the family Nannoceratopsiaceae. It is a genus related with Marine deposits. |
|
|
Scrinocassis[9] |
|
|
Miospores |
Dubious Genus with affinities with Scriniocassiaceae. Brackish Green Algae, related to lagoonar water bodies |
|
|
Porcellispora[9] |
|
|
Miospores |
Dubious Genus with affinities with Bryophyta |
|
|
Aratrisporites[9] |
|
|
Miospores |
Affinities with Isoetaceae. This genus is usually found in situ found related to genera such as Cyclostrobus, Lycostrobus and Annalepis. |
|
|
Foveosporites[9] |
|
|
Miospores |
Affinities with the family Lycopodiaceae inside Lycopodiopsida. Lycopod spores, related with herbaceous to arbustive flora common on humid environments |
 Extant Lycopodium specimens. Genera like Foveosporites probably come from a similar plant |
|
Densoisporites[9] |
|
|
Miospores |
Affinities with the Selaginellaceae inside Lycopsida. Herbaceous Lycophyte flora, similar to ferns, related to humid settings. |
 Extant Selaginella, typical example of Selaginellaceae. Genera like Densoisporites probably came from a similar or related plant |
|
Calamospora[9] |
|
|
Miospores |
Affinities with the Calamitaceae inside Equisetales. Horsetails, herbaceous flora related to high humid environments, flooding tolerant plants. |
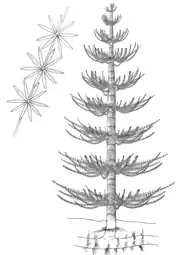 Recosntruction of the Genus Calamites, found associated with Calamospora |
|
Styxisporites[9] |
|
|
Miospores |
Uncertain Fern Miospores whose affinity cannot be concreted beyond Pteridophytes. |
|
|
Ischyosporites[9] |
|
|
Miospores | ||
|
Verrucosisporites[9] |
|
|
Miospores | ||
|
Deltoidospora[9] |
|
|
Miospores | ||
|
Skarbysporites[9] |
|
|
Miospores | ||
|
Murospora[9] |
|
|
Miospores | ||
|
Klukisporites[9] |
|
|
Miospores |
Affinities with the family Lygodiaceae inside Polypodiopsida. Climbing fern spores |
.gif) Example of extant Lygodium, Klukisporites come probably from similar genera or maybe a species from the genus |
|
Monolites[9] |
|
|
Miospores |
Affinities with the Polypodiaceae inside Polypodiales. Ferns of several sizes, from both dry land and near water environments. |
 Example of extant Drynaria specimens, Leptolepidites come probably from similar genera |
|
Leptolepidites[9] |
|
|
Miospores |
Affinities with the family Dennstaedtiaceae inside Polypodiales. Forested areas Fern Spores |
.jpg.webp) Example of extant Dennstaedtia specimens, Leptolepidites come probably from similar genera |
|
Granulatisporites[9] |
|
|
Miospores |
Affinities with Dipteridaceae inside Pteridophyta. Fern spores related to freshwater ponds. |
_(7113325651).jpg.webp) Example of extant Dipteris specimens, Granulatisporites come probably from similar genera |
|
Duplexisporites[9] |
|
|
Miospores |
Affinities with the family Cyatheaceae inside Cyatheales. Arboreal Fern Spores |
 Example of extant Cyathea, Duplexisporites come probably from similar genera |
|
Obtusisporites[9] |
|
|
Miospores | ||
|
Concavisporites[9] |
|
|
Miospores |
Affinities with the genus Gleichenia inside Gleicheniaceae. Tropical Ferns related to humid ferric soils. |
 Example of extant Gleichenia, Concavisporites come probably from similar genera |
|
Matonisporites[9] |
|
|
Miospores |
Affinities with the genus Matoniaceae inside Gleicheniales. Ferns of several sizes, from both dry land and near water environments. |
|
|
Vitreisporites[9] |
|
|
Pollen |
Pollen from the Family Caytoniaceae inside Caytoniales. Caytoniaceae are a complex group of Mesozoic Fossil floras, that can be related to both Peltaspermales and Ginkgoaceae. |
|
|
Cycadopites[9] |
|
|
Pollen |
Affinities with the family Cycadaceae inside Cycadales. |
 Extant Cycas platyphylla. Cycadopites maybe come from a related plant |
|
Bennettiteaepollenites[9] |
|
|
Pollen |
Affinities with Bennettitaceae inside Bennettitales. Abundant and Dry environment indicator |
|
|
Callialasporites[9] |
|
|
Pollen |
Affinities with the family Araucariaceae inside Pinales. Conifer pollen from medium to large arboreal plants |
 Extant Araucaria. Callialasporites maybe came from a related plant |
|
|
Pollen |
Affinities with the Hirmeriellaceae inside Pinopsida. Dominant Palynological residue, indicator of Dry conditions |
||
|
Podocarpidites[9] |
|
|
Pollen |
Affinities with the Podocarpaceae inside Pinopsida. Conifer pollen from medium to large arboreal plants |
.jpg.webp) Extant Podocarpus. Podocarpidites maybe come from a related plant |
|
Cerebropollenites[9] |
|
|
Pollen |
Affinities with both Sciadopityaceae and Miroviaceae inside Pinopsida. This Pollen resemblance with extant Sciadopitys suggest that Miroviaceae can be an extinct lineage of sciadopityaceaous-like plants.[14] |
 Extant Sciadopitys. Cerebropollenites likely come from a related plant |
|
Pityosporites[9] |
|
|
Pollen |
Affinities with the family Pinaceae inside Pinopsida. Conifer pollen from medium to large arboreal plants |
 Extant Tsuga Cone, example of the Abietoideae. Pinaceae Pollen like Pityosporites likely come from a similar plant |
Plant remains
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Isolated Stems |
Affinities with Equisetaceae inside Sphenopsida. Near water plants, associated with static freshwater ponds and other humid environments. |
 Example of Equisetites specimen | |
|
|
Isolated pinnae |
A Fern of the family Polypodiales inside Polypodiidae. Common cosmopolitan Mesozoic fern genus. Recent research has reinterpreted it a stem group of the Polypodiales (Closely related with the extant genera Dennstaedtia, Lindsaea, and Odontosoria).[15] |
.jpg.webp) Coniopteris specimen | |
|
|
Isolated pinnae |
Affinities with Umkomasiaceae inside Peltaspermopsida. Large Tree ferns associated to humid conditions. The dominant floral remain over the mangrove-type layers |
||
|
|
Pollen Organs |
Affinities with Caytoniales inside Peltaspermopsida. Reproductive organ of the Peltaspermales, with berry like cupules with numerous small seeds arrayed along axes. |
||
|
|
Isolated leaflets |
Affinities with Williamsoniaceae inside Bennettitales. Cycadales-Like medium sized trees. The most abundant flora on the Budos Mountain Limestone. |
.jpg.webp) Otozamites specimen | |
|
|
Isolated leaflets |
Affinities with Williamsoniaceae inside Bennettitales. |
 Ptilophyllum specimen | |
|
|
Isolated leaflets |
Affinities with Williamsoniaceae inside Bennettitales. |
 Zamites specimen | |
|
|
Branched shoots |
Affinities with Ginkgoales inside Ginkgoopsida. Ginko Relatives with a more wider leaf, representing among the best specimens found on the mediterranean realm. |
||
|
|
Branched shoots |
Affinities with Araucariaceae or Hirmeriellaceae inside Pinales. |
_(cropped).jpg.webp) Example of Brachyphyllum specimen | |
|
|
Branched shoots |
Affinities with Araucariaceae or Hirmeriellaceae inside Pinales. |
 Example of Pagiophyllum specimen | |
|
|
Branched shoots |
Affinities with Krassiloviaceae inside Voltziales. |
||
|
|
Branched shoots |
Affinities with Cupressaceae inside Coniferales. Leaves from Arbustive to Arboreal Coniferous trees. |
_(Judith_River_Group%252C_Upper_Cretaceous%253B_Montana_or_Canada)_(25210755717).jpg.webp) Elatides specimens | |
Bivalves
The Budos Mountain facies, like the Rotzo Formation, is known mostly due to its massive bivalve associations of the genera Lithiotis, Cochlearites and Lithioperna that extended all along the Pliensbachian-Toarcian Adriatic-Dinaric-Hellenic Platforms forming mass accumulations of specimens that formed Reef-Like structures.[16] This fauna appeared after the early Pliensbachian C-cycle perturbation, that triggered the diffusion of the Lithiotis Fauna, noted on the rapid widespread of this biota after the event layers.[16] All of the genera related with this fauna appeared on the lower Jurassic, and all but one became extinct before the Middle Jurassic.[17] This "Reefs" had a strong zonation, starting with the bivalves Gervilleioperna and Mytiloperna, restricted to intertidal and shallow-subtidal facies. Lithioperna is limited to lagoonal subtidal facies and even in some low-oxygen environments. Finally Lithiotis and Cochlearites are found in subtidal facies, constructing buildups.[17] This sections formed various kinds of ecosystems on the Trento platform, where it appeared in branched corals filled with (Spongiomorpha), Domal corals (Stromatoporida), tubular corals, Styllophyllidae corals, unidentified Cerioidea colonial corals, regular echinoid debris, sponges, and the solitary coral Opelismilia sp., with also aggregated snail shells.[17]
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
Cochlearites[1] |
|
|
|
A clam, incertae sedis inside Pterioida. A large bivalve, with a subequivalved shell, up to 60–70 cm high. It is one of the Three main bivalves recovered on the Lithiotis Facies, with its accumulations generally overlying megalodontid coquinas. |
|
|
Lithioperna[1] |
|
|
|
A clam, incertae sedis inside Pterioida. This genus was found to be a bivalve with a byssate juvenile stage that developed different modes of life in adulthood depending on the individual density and bottom firmness. |
|
|
Lithiotis[1] |
|
|
|
An oyster, member of Malleidae inside Ostreida. It is the major Bivalve identified on the formation, and the genus that gives the name to the Lithiotis fauna. Large, large and aberrant bivalves present on mostly of the Trento Platform. Its accumulation have had different denominations on literature, such as banks, bioherms, biostromes, bivalve reefs or bivalve mounds. |
|
|
Gervilleioperna[1] |
|
|
Isolated Shells |
An Oyster, member of Malleidae inside Ostreida. On the Rotzo formation this genus become abundant along rootlets, indicative of a very shallow and restricted lagoon or marsh environment. |
|
|
Mytiloperna[1] |
|
|
Isolated Shells |
||
See also
- List of fossiliferous stratigraphic units in Italy
- Toarcian turnover
- Toarcian formations
- Marne di Monte Serrone, Italy
- Calcare di Sogno, Italy
- Posidonia Shale, Lagerstätte in Germany
- Ciechocinek Formation, Germany and Poland
- Krempachy Marl Formation, Poland & Slovakia
- Lava Formation, Lithuania
- Azilal Group, North Africa
- Whitby Mudstone, England
- Fernie Formation, Alberta and British Columbia
- Poker Chip Shale
- Whiteaves Formation, British Columbia
- Navajo Sandstone, Utah
- Los Molles Formation, Argentina
- Mawson Formation, Antarctica
- Kandreho Formation, Madagascar
- Kota Formation, India
- Cattamarra Coal Measures, Australia
References
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 Pantić, N. K. (1952). "Liassic flora from Budos mountain - Montenegro". Glasnik Prir. Muzeja SRP. Zem. 5 (1): 293–308.
- ↑ Mirkovic, M.; Kalezic, M.; Pajovic, M. (1977). "Osnovna geološka karta SFRJ 1: 100.000List Bar K34–63 (Basic Geologic Map of SFRY 1: 100.000–the Bar sheet)". Savezni geol. Zavod Beograd Zavod geol. Istraž. Crne Gore (2): 1962–1968.
- ↑ Pantic, N.; Grubic, A.; Sladic-Trifunovic, M. (1983). "The importance of Mesozoic floras and faunas from intraoceanic carbonate platforms for the interpretation of paleogeographic and geodynamic events in the Tethys". Boll. Soc. Pal. Ltaliana. 22 (2): 5–14.
- 1 2 Dragičević, I.; Velić, I. (2002). "The northeastern margin of the Adriatic Carbonate Platform". Geologia Croatica. 55 (2): 185–232. doi:10.4154/GC.2002.16. S2CID 73612045. Retrieved 28 January 2022.
- 1 2 3 Pantic, N. (1980). "Environments, Paleogeogeography and Tectonics". SGD Records of 1979. 21 (4): 7–13.
- 1 2 3 Čadjenović, D.; Kilibarda, Z.; Radulović, N. (2008). "Late Triassic to Late Jurassic evolution of the Adriatic carbonate platform and Budva Basin, southern Montenegro". Sedimentary Geology. 204 (2): 1–17. doi:10.1016/j.sedgeo.2007.12.005. Retrieved 28 January 2022.
- ↑ Crne, A.; Gorican, S.; Cadjenovic, D. (2006). "Lower Jurassic carbonate platform-to-basin transition at Mt. Rumija (Montenegro)". Volumina Jurassica. 4 (4): 82–83. Retrieved 28 January 2022.
- ↑ Vakhrameev, V. A. (1991). Jurassic and Cretaceous floras and climates of the Earth (PDF). Cambridge: Cambridge University Press. p. 21. Retrieved 28 January 2022.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 Pantić, N. K. (1981). "Macroflora and palynomorphs from Lower jurassic of Budos Mountain, Montenegro". Ann. Geol. Peninsule Balk. 45 (1): 157–171.
- 1 2 3 4 Pantić, N.K.; Duuc, S. (1990). "Palaeophytogeography of Jurassic land flores in Tethyan regions and its margins". Geol. An. Balk. Pol. 2 (1): 237–247.
- ↑ Barrón, E.; Ureta, S.; Goy, A.; Lassaletta, L. (2010). "Palynology of the Toarcian–Aalenian Global Boundary Stratotype Section and Point (GSSP) at Fuentelsaz (Lower–Middle Jurassic, Iberian Range, Spain)". Review of Palaeobotany and Palynology. 162 (1): 11–28. doi:10.1016/j.revpalbo.2010.04.003. Retrieved 28 January 2022.
- ↑ Barbacka, M.; Krobicki, M.; Iwańczuk, J.; Muceku, B. (2019). "Kora Jura időszaki növényi és Lithiotis-típusú kagylómaradványok az Albán Alpokban [The Early Jurassic association of plant remains and Lithiotis-type bivalves in the Albanian Alps]" (PDF). Annales Musei historico-naturalis hungarici. 111 (2): 103–114. Retrieved 28 January 2022.
- ↑ Krobicki, M. (2019). "Is there any connection between the Early Jurassic (Pliensbachian) Lithiotis-type bivalve facies of mangrove-type environments (Albanian Alps) and the origin of primitive crabs (Decapoda, Brachyura)?" (PDF). 20th Czech-Polish-Slovak Palaeontological Conference. 20 (2): 33. Retrieved 28 January 2022.
- ↑ Hofmann, Christa-Ch.; Odgerel, Nyamsambuu; Seyfullah, Leyla J. (2021). "The occurrence of pollen of Sciadopityaceae Luerss. through time". Fossil Imprint. 77 (2): 271–281. doi:10.37520/fi.2021.019. S2CID 245555379. Retrieved 27 December 2021.
- ↑ Li, Chunxiang; Miao, Xinyuan; Zhang, Li-Bing; Ma, Junye; Hao, Jiasheng (January 2020). "Re-evaluation of the systematic position of the Jurassic–Early Cretaceous fern genus Coniopteris". Cretaceous Research. 105: 104136. doi:10.1016/j.cretres.2019.04.007. S2CID 146355798.
- 1 2 Franceschi, M.; Dal Corso, J.; Posenato, R.; Roghi, G.; Masetti, D.; Jenkyns, H.C. (2014). "Early Pliensbachian (Early Jurassic) C-isotope perturbation and the diffusion of the Lithiotis Fauna: Insights from the western Tethys". Palaeogeography, Palaeoclimatology, Palaeoecology. 410 (4): 255–263. Bibcode:2014PPP...410..255F. doi:10.1016/j.palaeo.2014.05.025. Retrieved 3 January 2022.
- 1 2 3 Fraser, N.M.; Bottjer, D.J.; Fischer, A.G. (2004). "Dissecting "Lithiotis" Bivalves: Implications for the Early Jurassic Reef Eclipse". PALAIOS. 19 (1): 51–67. Bibcode:2004Palai..19...51F. doi:10.1669/0883-1351(2004)019<0051:DLBIFT>2.0.CO;2. S2CID 128632794. Retrieved 3 January 2022.