
In biochemistry, biomolecular condensates are a class of membrane-less organelles and organelle subdomains, which carry out specialized functions within the cell. Unlike many organelles, biomolecular condensate composition is not controlled by a bounding membrane. Instead, condensates can form and maintain organization through a range of different processes, the most well-known of which is phase separation of proteins, RNA and other biopolymers into either colloidal emulsions, gels, liquid crystals, solid crystals or aggregates within cells.[1]
History
Micellar theory

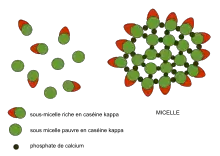
The micellar theory of Carl Nägeli was developed from his detailed study of starch granules in 1858.[2] Amorphous substances such as starch and cellulose were proposed to consist of building blocks, packed in a loosely crystalline array to form what he later termed “micelles”. Water could penetrate between the micelles, and new micelles could form in the interstices between old micelles. The swelling of starch grains and their growth was described by a molecular-aggregate model, which he also applied to the cellulose of the plant cell wall. The modern usage of 'micelle' refers strictly to lipids, but its original usage clearly extended to other types of biomolecule, and this legacy is reflected to this day in the description of milk as being composed of 'casein micelles'.
Colloidal phase separation theory
.tif.jpg.webp)
The concept of intracellular colloids as an organizing principle for the compartmentalization of living cells dates back to the end of the 19th century, beginning with William Bate Hardy and Edmund Beecher Wilson who described the cytoplasm (then called 'protoplasm') as a colloid.[3][4] Around the same time, Thomas Harrison Montgomery Jr. described the morphology of the nucleolus, an organelle within the nucleus, which has subsequently been shown to form through intracellular phase separation.[5] WB Hardy linked formation of biological colloids with phase separation in his study of globulins, stating that: "The globulin is dispersed in the solvent as particles which are the colloid particles and which are so large as to form an internal phase",[6] and further contributed to the basic physical description of oil-water phase separation.[7]
Colloidal phase separation as a driving force in cellular organisation appealed strongly to Stephane Leduc, who wrote in his influential 1911 book The Mechanism of Life: "Hence the study of life may be best begun by the study of those physico-chemical phenomena which result from the contact of two different liquids. Biology is thus but a branch of the physico-chemistry of liquids; it includes the study of electrolytic and colloidal solutions, and of the molecular forces brought into play by solution, osmosis, diffusion, cohesion, and crystallization."[8]
The primordial soup theory of the origin of life, proposed by Alexander Oparin in Russian in 1924 (published in English in 1936)[9] and by J.B.S. Haldane in 1929,[10] suggested that life was preceded by the formation of what Haldane called a "hot dilute soup" of "colloidal organic substances", and which Oparin referred to as 'coacervates' (after de Jong[11]) – particles composed of two or more colloids which might be protein, lipid or nucleic acid. These ideas strongly influenced the subsequent work of Sidney W. Fox on proteinoid microspheres.
Support from other disciplines
When cell biologists largely abandoned colloidal phase separation, it was left to relative outsiders – agricultural scientists and physicists – to make further progress in the study of phase separating biomolecules in cells.
Beginning in the early 1970s, Harold M Farrell Jr. at the US Department of Agriculture developed a colloidal phase separation model for milk casein micelles that form within mammary gland cells before secretion as milk.[12]
Also in the 1970s, physicists Tanaka & Benedek at MIT identified phase-separation behaviour of gamma-crystallin proteins from lens epithelial cells and cataracts in solution,[13][14][15][16][17] which Benedek referred to as 'protein condensation'.[18]
_(14763034584).jpg.webp)
In the 1980s and 1990s, Athene Donald's polymer physics lab in Cambridge extensively characterised phase transitions / phase separation of starch granules from the cytoplasm of plant cells, which behave as liquid crystals.[19][20][21][22][23][24][25][26]
In 1991, Pierre-Gilles de Gennes received the Nobel Prize in Physics for developing a generalized theory of phase transitions with particular applications to describing ordering and phase transitions in polymers.[27] Unfortunately, de Gennes wrote in Nature that polymers should be distinguished from other types of colloids, even though they can display similar clustering and phase separation behaviour,[28] a stance that has been reflected in the reduced usage of the term colloid to describe the higher-order association behaviour of biopolymers in modern cell biology and molecular self-assembly.
Phase separation revisited
Advances in confocal microscopy at the end of the 20th century identified proteins, RNA or carbohydrates localising to many non-membrane bound cellular compartments within the cytoplasm or nucleus which were variously referred to as 'puncta/dots',[29][30][31][32] 'signalosomes',[33][34] 'granules',[35] 'bodies', 'assemblies',[32] 'paraspeckles', 'purinosomes',[36] 'inclusions', 'aggregates' or 'factories'. During this time period (1995-2008) the concept of phase separation was re-borrowed from colloidal chemistry & polymer physics and proposed to underlie both cytoplasmic and nuclear compartmentalization.[37][38][39][40][41][42][43][44][45][46]
Since 2009, further evidence for biomacromolecules undergoing intracellular phase transitions (phase separation) has been observed in many different contexts, both within cells and in reconstituted in vitro experiments.[47][48][49][50][51][52][53]
The newly coined term "biomolecular condensate"[54] refers to biological polymers (as opposed to synthetic polymers) that undergo self assembly via clustering to increase the local concentration of the assembling components, and is analogous to the physical definition of condensation.[55][54]
In physics, condensation typically refers to a gas–liquid phase transition.
In biology the term 'condensation' is used much more broadly and can also refer to liquid–liquid phase separation to form colloidal emulsions or liquid crystals within cells, and liquid–solid phase separation to form gels,[1] sols, or suspensions within cells as well as liquid-to-solid phase transitions such as DNA condensation during prophase of the cell cycle or protein condensation of crystallins in cataracts.[18] With this in mind, the term 'biomolecular condensates' was deliberately introduced to reflect this breadth (see below). Since biomolecular condensation generally involves oligomeric or polymeric interactions between an indefinite number of components, it is generally considered distinct from formation of smaller stoichiometric protein complexes with defined numbers of subunits, such as viral capsids or the proteasome – although both are examples of spontaneous molecular self-assembly or self-organisation.
Mechanistically, it appears that the conformational landscape[56] (in particular, whether it is enriched in extended disordered states) and multivalent interactions between intrinsically disordered proteins (including cross-beta polymerisation),[57] and/or protein domains that induce head-to-tail oligomeric or polymeric clustering,[58] might play a role in phase separation of proteins.
Examples

Many examples of biomolecular condensates have been characterized in the cytoplasm and the nucleus that are thought to arise by either liquid–liquid or liquid–solid phase separation.
Cytoplasmic condensates
- Lewy bodies
- Stress granule
- P-body
- Germline P-granules – oskar
- Starch granules
- Glycogen granules[59]
- Frodosomes (Dact1)[60]
- Corneal lens formation and cataracts[13][14][15][16][17][18]
- Other cytoplasmic inclusions such as pigment granules or cytoplasmic crystals
- Purinosomes[36]
- Misfolded protein aggregation such as amyloid fibrils or mutant Haemoglobin S (HbS) fibres in sickle cell disease
- Signalosomes, such as the supramolecular assemblies in the Wnt signaling pathway.[61][62]
- It can also be argued that cytoskeletal filaments form by a polymerisation process similar to phase separation, except ordered into filamentous networks instead of amorphous droplets or granules.
- Bacteria Ribonucleoprotein Bodies (BR-bodies)- In recent studies it has been shown that bacteria RNA degradosomes can assemble into phase‐separated structures, termed bacterial ribonucleoprotein bodies (BR‐bodies), with many analogous properties to eukaryotic processing bodies and stress granules.[63]
- FLOE1 granules: FLOE1 is a prion-like seed-specific protein that controls plant seed germination via phase separation into biomolecular condensates.[64]
Nuclear condensates
Other nuclear structures including heterochromatin form by mechanisms similar to phase separation, so can also be classified as biomolecular condensates.
Plasma membrane associated condensates
- Membrane protein, or membrane-associated protein, clustering at neurological synapses, cell-cell tight junctions, or other membrane domains.[65]
Secreted extracellular condensates
- Secreted thyroglobulin colloid and colloid nodules of the thyroid gland
- Secreted casein ‘micelles’ of the mammary gland
- Serum albumin and globulins
- Secreted lysozyme[66][42]
Lipid-enclosed organelles and lipoproteins are not considered condensates
Typical organelles or endosomes enclosed by a lipid bilayer are not considered biomolecular condensates. In addition, lipid droplets are surrounded by a lipid monolayer in the cytoplasm, or in milk, or in tears,[67] so appear to fall under the 'membrane bound' category. Finally, secreted LDL and HDL lipoprotein particles are also enclosed by a lipid monolayer. The formation of these structures involves phase separation to from colloidal micelles or liquid crystal bilayers, but they are not classified as biomolecular condensates, as this term is reserved for non-membrane bound organelles.
Liquid–liquid phase separation (LLPS) in biology

Liquid biomolecular condensates
Liquid–liquid phase separation (LLPS) generates a subtype of colloid known as an emulsion that can coalesce to from large droplets within a liquid. Ordering of molecules during liquid–liquid phase separation can generate liquid crystals rather than emulsions. In cells, LLPS produces a liquid subclass of biomolecular condensate that can behave as either an emulsion or liquid crystal.
The term biomolecular condensates was introduced in the context of intracellular assemblies as a convenient and non-exclusionary term to describe non-stoichiometric assemblies of biomolecules.[54] The choice of language here is specific and important. It has been proposed that many biomolecular condensates form through liquid–liquid phase separation (LLPS) to form colloidal emulsions or liquid crystals in living organisms, as opposed to liquid–solid phase separation to form crystals/aggregates in gels,[1] sols or suspensions within cells or extracellular secretions.[68] However, unequivocally demonstrating that a cellular body forms through liquid–liquid phase separation is challenging,[69][47][70][71] because different material states (liquid vs. gel vs. solid) are not always easy to distinguish in living cells.[72][73] The term "biomolecular condensate" directly addresses this challenge by making no assumption regarding either the physical mechanism through which assembly is achieved, nor the material state of the resulting assembly. Consequently, cellular bodies that form through liquid–liquid phase separation are a subset of biomolecular condensates, as are those where the physical origins of assembly are unknown. Historically, many cellular non-membrane bound compartments identified microscopically fall under the broad umbrella of biomolecular condensates.
In physics, phase separation can be classified into the following types of colloid, of which biomolecular condensates are one example:
| Medium/phase | Dispersed phase | |||
|---|---|---|---|---|
| Gas | Liquid | Solid | ||
| Dispersion medium |
Gas | No such colloids are known. Helium and xenon are known to be immiscible under certain conditions.[74][75] |
Liquid aerosol Examples: fog, clouds, condensation, mist, hair sprays | Solid aerosol Examples: smoke, ice cloud, atmospheric particulate matter |
| Liquid | Foam Example: whipped cream, shaving cream, Gas vesicles | Emulsion or Liquid crystal Examples: milk, mayonnaise, hand cream, latex, biological membranes, micelles, lipoproteins, silk, liquid biomolecular condensates | Sol or suspension Examples: pigmented ink, sediment, precipitates, aggregates, fibres/fibrils/filaments, crystals, solid biomolecular condensates | |
| Solid | Solid foam Examples: aerogel, styrofoam, pumice | Gel Examples: agar, gelatin, jelly, gel-like biomolecular condensates | Solid sol Example: cranberry glass | |
In biology, the most relevant forms of phase separation are either liquid–liquid or liquid–solid, although there have been reports of gas vesicles surrounded by a phase separated protein coat in the cytoplasm of some microorganisms.[76]
Wnt signalling
One of the first discovered examples of a highly dynamic intracellular liquid biomolecular condensate with a clear physiological function were the supramolecular complexes (Wnt signalosomes) formed by components of the Wnt signaling pathway.[44][61][62] The Dishevelled (Dsh or Dvl) protein undergoes clustering in the cytoplasm via its DIX domain, which mediates protein clustering (polymerisation) and phase separation, and is important for signal transduction.[29][30][31][32][34][44] The Dsh protein functions both in planar polarity and Wnt signalling, where it recruits another supramolecular complex (the Axin complex) to Wnt receptors at the plasma membrane. The formation of these Dishevelled and Axin containing droplets is conserved across metazoans, including in Drosophila, Xenopus, and human cells.
P granules
Another example of liquid droplets in cells are the germline P granules in Caenorhabditis elegans.[68][47] These granules separate out from the cytoplasm and form droplets, as oil does from water. Both the granules and the surrounding cytoplasm are liquid in the sense that they flow in response to forces, and two of the granules can coalesce when they come in contact. When (some of) the molecules in the granules are studied (via fluorescence recovery after photobleaching), they are found to rapidly turnover in the droplets, meaning that molecules diffuse into and out of the granules, just as expected in a liquid droplet. The droplets can also grow to be many molecules across (micrometres)[47] Studies of droplets of the Caenorhabditis elegans protein LAF-1 in vitro[77] also show liquid-like behaviour, with an apparent viscosity Pa s. This is about a ten thousand times that of water at room temperature, but it is small enough to enable the LAF-1 droplets to flow like a liquid. Generally, interaction strength (affinity)[78] and valence (number of binding sites)[53] of the phase separating biomolecules influence their condensates viscosity, as well as their overall tendency to phase separate.

Liquid–liquid phase separation in human disease
Growing evidence suggests that anomalies in biomolecular condensates formation can lead to a number of human pathologies[79] such as cancer and neurodegenerative diseases.[80][81]
Synthetic biomolecular condensates
Biomolecular condensates can be synthesized for a number of purposes. Synthetic biomolecular condensates are inspired by endogenous biomolecular condensates, such as nucleoli, P bodies, and stress granules, which are essential to normal cellular organization and function.[82][83]
Synthetic condensates are an important tool in synthetic biology, and have a wide and growing range of applications. Engineered synthetic condensates allow for probing cellular organization, and enable the creation of novel functionalized biological materials, which have the potential to serve as drug delivery platforms and therapeutic agents.[84]
Design and control
Despite the dynamic nature and lack of binding specificity that govern the formation of biomolecular condensates, synthetic condensates can still be engineered to exhibit different behaviors. One popular way to conceptualize condensate interactions and aid in design is through the "sticker-spacer" framework.[85] Multivalent interaction sites, or "stickers", are separated by "spacers", which provide the conformational flexibility and physically separate individual interaction modules from one another. Proteins regions identified as ‘stickers’ usually consist of Intrinsically Disordered Regions (IDRs) that act as "sticky" biopolymers via short patches of interacting residues patterned along their unstructured chain, which collectively promote LLPS.[86] By modifying the sticker-spacer framework, i.e. the polypeptide and RNA sequences as well as their mixture compositions, the material properties (viscous and elastic regimes) of condensates can be tuned to design novel condensates.[87]
Other tools outside of tuning the sticker-spacer framework can be used to give new functionality and to allow for high temporal and spatial control over synthetic condensates. One way to gain temporal control over the formation and dissolution of biomolecular condensates is by using optogenetic tools. Several different systems have been developed which allow for control of condensate formation and dissolution which rely on chimeric protein expression, and light or small molecule activation.[88] In one system,[89] proteins are expressed in a cell which contain light-activated oligomerization domains fused to IDRs. Upon irradiation with a specific wavelength of light, the oligomerization domains bind each other and form a ‘core’, which also brings multiple IDRs close together because they are fused to the oligomerization domains. The recruitment of multiple IDRs effectively creates a new biopolymer with increased valency. This increased valency allows for the IDRs to form multivalent interactions and trigger LLPS. When the activation light is stopped, the oligomerization domains disassemble, causing the dissolution of the condensate. A similar system[90] achieves the same temporal control of condensate formation by using light-sensitive ‘caged’ dimerizers. In this case, light-activation removes the dimerizer cage, allowing it to recruit IDRs to multivalent cores, which then triggers phase separation. Light-activation of a different wavelength results in the dimerizer being cleaved, which then releases the IDRs from the core and consequentially dissolves the condensate. This dimerizer system requires significantly reduced amounts of laser light to operate, which is advantageous because high intensity light can be toxic to cells.
Optogenetic systems can also be modified to gain spatial control over the formation of condensates. Multiple approaches have been developed to do so. In one approach,[91] which localizes condensates to specific genomic regions, core proteins are fused to proteins such as TRF1 or catalytically dead Cas9, which bind specific genomic loci. When oligomerization is trigger by light activation, phase separation is preferentially induced on the specific genomic region which is recognized by fusion protein. Because condensates of the same composition can interact and fuse with each other, if they are tethered to specific regions of the genome, condensates can be used to alter the spatial organization of the genome, which can have effects on gene expression.[91]
As biochemical reactors
Synthetic condensates offer a way to probe cellular function and organization with high spatial and temporal control, but can also be used to modify or add functionality to the cell. One way this is accomplished is by modifying the condensate networks to include binding sites for other proteins of interest, thus allowing the condensate to serve as a scaffold for protein release or recruitment.[92] These binding sites can be modified to be sensitive to light activation or small molecule addition, thus giving temporal control over the recruitment of a specific protein of interest. By recruiting specific proteins to condensates, reactants can be concentrated to increase reaction rates or sequestered to inhibit reactivity.[93] In addition to protein recruitment, condensates can also be designed which release proteins in response to certain stimuli. In this case, a protein of interest can be fused to a scaffold protein via a photocleavable linker. Upon irradiation, the linker is broken, and the protein is released from the condensate. Using these design principles, proteins can either be released to, or sequestered from, their native environment, allowing condensates to serve as a tool to alter the biochemical activity of specific proteins with a high level of control.[92]
Methods to study condensates
A number of experimental and computational methods have been developed to examine the physico-chemical properties and underlying molecular interactions of biomolecular condensates. Experimental approaches include phase separation assays using bright-field imaging or fluorescence microscopy, as well as fluorescence recovery after photobleaching (FRAP).[94] Computational approaches include coarse-grained molecular dynamics simulations and circuit topology analysis.[95]
Coarse-grained molecular models
Molecular dynamics and Monte Carlo simulations have been extensively used to gain insights into the formation and the material properties of biomolecular condensates.[96] Although molecular models of different resolution have been employed,[97][98][99] modelling efforts have mainly focused on coarse-grained models of intrinsically disordered proteins, wherein amino acid residues are represented by single interaction sites. Compared to more detailed molecular descriptions, residue-level models provide high computational efficiency, which enables simulations to cover the long length and time scales required to study phase separation. Moreover, the resolution of these models is sufficiently detailed to capture the dependence on amino acid sequence of the properties of the system.[96]
Several residue-level models of intrinsically disordered proteins have been developed in recent years. Their common features are (i) the absence of an explicit representation of solvent molecules and salt ions, (ii) a mean-field description of the electrostatic interactions between charged residues (see Debye–Hückel theory), and (iii) a set of "stickiness" parameters which quantify the strength of the attraction between pairs of amino acids. In the development of most residue-level models, the stickiness parameters have been derived from hydrophobicity scales[100] or from a bioinformatic analysis of crystal structures of folded proteins.[101][102] Further refinement of the parameters has been achieved through iterative procedures which maximize the agreement between model predictions and a set of experiments,[103][104][105][106][107][108] or by leveraging data obtained from all-atom molecular dynamics simulations.[102]
Residue-level models of intrinsically disordered proteins have been validated by direct comparison with experimental data, and their predictions have been shown to be accurate across diverse amino acid sequences.[103][104][105][102][107][109][108] Examples of experimental data used to validate the models are radii of gyration of isolated chains and saturation concentrations, which are threshold protein concentrations above which phase separation is observed.[110]
Although intrinsically disordered proteins often play important roles in condensate formation,[111] many biomolecular condensates contain multi-domain proteins constituted by folded domains connected by intrinsically disordered regions.[112] Current residue-level models are only applicable to the study of condensates of intrinsically disordered proteins and nucleic acids.[113][102][114][115][116][108] Including an accurate description of the folded domains in these models will considerably widen their applicability.[117][96]
References
- 1 2 3 Garaizar, Adiran; Espinosa, Jorge R.; Joseph, Jerelle A.; Collepardo-Guevara, Rosana (2022-03-15). "Kinetic interplay between droplet maturation and coalescence modulates shape of aged protein condensates". Scientific Reports. 12 (1): 4390. Bibcode:2022NatSR..12.4390G. doi:10.1038/s41598-022-08130-2. ISSN 2045-2322. PMC 8924231. PMID 35293386.
- ↑ Farlow, William G. (1890). "Proceedings of the American Academy of Arts and Sciences". 26. American Academy of Arts & Sciences: 376–381. JSTOR 20013496.
{{cite journal}}: Cite journal requires|journal=(help) - ↑ Wilson EB (July 1899). "The Structure of Protoplasm". Science. 10 (237): 33–45. Bibcode:1899Sci....10...33W. doi:10.1126/science.10.237.33. PMID 17829686.
- ↑ Hardy WB (May 1899). "On the structure of cell protoplasm: Part I. The Structure produced in a Cell by Fixative and Post-mortem change. The Structure of Colloidal matter and the Mechanism of Setting and of Coagulation". The Journal of Physiology. 24 (2): 158–210.1. doi:10.1113/jphysiol.1899.sp000755. PMC 1516635. PMID 16992486.
- ↑ Montgomery T (1898). "Comparative cytological studies, with especial regard to the morphology of the nucleolus". Journal of Morphology. 15 (1): 265–582. doi:10.1002/jmor.1050150204. S2CID 84531494.
- ↑ Hardy WB (December 1905). "Colloidal solution. The globulins". The Journal of Physiology. 33 (4–5): 251–337. doi:10.1113/jphysiol.1905.sp001126. PMC 1465795. PMID 16992817.
- ↑ Hardy WB (1912). "The tension of composite fluid surfaces and the mechanical stability of films of fluid". Proceedings of the Royal Society A. 86 (591): 610–635. Bibcode:1912RSPSA..86..610H. doi:10.1098/rspa.1912.0053.
- ↑ Leduc S (1911). "The Mechanism of Life".
- ↑ Oparin A. "The Origin of Life" (PDF).
- ↑ Haldane JB. "The Origin of Life" (PDF).
- ↑ Bungenberg de Jong HG, Kruyt HR. "Coacervation (partial miscibility in colloid systems". Proc. K. Ned. Akad. Wet 1929. 32: 849–856.
- ↑ Farrell HM (September 1973). "Models for casein micelle formation". Journal of Dairy Science. 56 (9): 1195–206. doi:10.3168/jds.S0022-0302(73)85335-4. PMID 4593735.
- 1 2 Tanaka T, Benedek GB (June 1975). "Observation of protein diffusivity in intact human and bovine lenses with application to cataract". Investigative Ophthalmology. 14 (6): 449–56. PMID 1132941.
- 1 2 Tanaka T, Ishimoto C, Chylack LT (September 1977). "Phase separation of a protein-water mixture in cold cataract in the young rat lens". Science. 197 (4307): 1010–2. Bibcode:1977Sci...197.1010T. doi:10.1126/science.887936. PMID 887936.
- 1 2 Ishimoto C, Goalwin PW, Sun ST, Nishio I, Tanaka T (September 1979). "Cytoplasmic phase separation in formation of galactosemic cataract in lenses of young rats". Proceedings of the National Academy of Sciences of the United States of America. 76 (9): 4414–6. Bibcode:1979PNAS...76.4414I. doi:10.1073/pnas.76.9.4414. PMC 411585. PMID 16592709.
- 1 2 Thomson JA, Schurtenberger P, Thurston GM, Benedek GB (October 1987). "Binary liquid phase separation and critical phenomena in a protein/water solution". Proceedings of the National Academy of Sciences of the United States of America. 84 (20): 7079–83. Bibcode:1987PNAS...84.7079T. doi:10.1073/pnas.84.20.7079. PMC 299233. PMID 3478681.
- 1 2 Broide ML, Berland CR, Pande J, Ogun OO, Benedek GB (July 1991). "Binary-liquid phase separation of lens protein solutions". Proceedings of the National Academy of Sciences of the United States of America. 88 (13): 5660–4. Bibcode:1991PNAS...88.5660B. doi:10.1073/pnas.88.13.5660. PMC 51937. PMID 2062844.
- 1 2 3 Benedek GB (September 1997). "Cataract as a protein condensation disease: the Proctor Lecture". Investigative Ophthalmology & Visual Science. 38 (10): 1911–21. PMID 9331254.
- ↑ Waigh TA, Gidley MJ, Komanshek BU, Donald AM (September 2000). "The phase transformations in starch during gelatinisation: a liquid crystalline approach". Carbohydrate Research. 328 (2): 165–76. doi:10.1016/s0008-6215(00)00098-7. PMID 11028784.
- ↑ Jenkins PJ, Donald AM (1998). "Gelatinisation of starch: A combined SAXS/WAXS/DSC and SANS study". Carbohydrate Research. 308 (1–2): 133. doi:10.1016/S0008-6215(98)00079-2.
- ↑ Jenkins PJ, Donald AM (December 1995). "The influence of amylose on starch granule structure". International Journal of Biological Macromolecules. 17 (6): 315–21. doi:10.1016/0141-8130(96)81838-1. PMID 8789332.
- ↑ Jenkins PJ, Cameron RE, Donald AM (1993). "A Universal Feature in the Structure of Starch Granules from Different Botanical Sources". Starch - Stärke. 45 (12): 417. doi:10.1002/star.19930451202.
- ↑ Donald AM, Windle AH, Hanna S (1993). "Liquid Crystalline Polymers". Physics Today. 46 (11): 87. Bibcode:1993PhT....46k..87D. doi:10.1063/1.2809100. hdl:2060/19900017655.
- ↑ Windle, A.H.; Donald, A.D. (1992). Liquid crystalline polymers. Cambridge, UK: Cambridge University Press. ISBN 978-0-521-30666-9.
- ↑ Starch: structure and functionality. Cambridge, England: Royal Society of Chemistry. 1997. ISBN 978-0-85404-742-0.
- ↑ The importance of polymer science for biological systems: University of York. Cambridge, England: Royal Society of Chemistry. March 2008. ISBN 978-0-85404-120-6.
- ↑ "The Nobel Prize in Physics 1991". /www.nobelprize.org.
- ↑ de Gennes PG (July 2001). "Ultradivided matter". Nature. 412 (6845): 385. Bibcode:2001Natur.412..385D. doi:10.1038/35086662. PMID 11473291. S2CID 39983702.
- 1 2 Cliffe A, Hamada F, Bienz M (May 2003). "A role of Dishevelled in relocating Axin to the plasma membrane during wingless signaling". Current Biology. 13 (11): 960–6. doi:10.1016/S0960-9822(03)00370-1. PMID 12781135. S2CID 15211115.
- 1 2 Schwarz-Romond T, Merrifield C, Nichols BJ, Bienz M (November 2005). "The Wnt signalling effector Dishevelled forms dynamic protein assemblies rather than stable associations with cytoplasmic vesicles". Journal of Cell Science. 118 (Pt 22): 5269–77. doi:10.1242/jcs.02646. PMID 16263762. S2CID 16988383.
- 1 2 Schwarz-Romond T, Fiedler M, Shibata N, Butler PJ, Kikuchi A, Higuchi Y, Bienz M (June 2007). "The DIX domain of Dishevelled confers Wnt signaling by dynamic polymerization". Nature Structural & Molecular Biology. 14 (6): 484–92. doi:10.1038/nsmb1247. PMID 17529994. S2CID 29584068.
- 1 2 3 Schwarz-Romond T, Metcalfe C, Bienz M (July 2007). "Dynamic recruitment of axin by Dishevelled protein assemblies". Journal of Cell Science. 120 (Pt 14): 2402–12. doi:10.1242/jcs.002956. PMID 17606995. S2CID 23270805.
- ↑ Bilic J, Huang YL, Davidson G, Zimmermann T, Cruciat CM, Bienz M, Niehrs C (June 2007). "Wnt induces LRP6 signalosomes and promotes dishevelled-dependent LRP6 phosphorylation". Science. 316 (5831): 1619–22. Bibcode:2007Sci...316.1619B. doi:10.1126/science.1137065. PMID 17569865. S2CID 25980578.
- 1 2 Bienz M (October 2014). "Signalosome assembly by domains undergoing dynamic head-to-tail polymerization". Trends in Biochemical Sciences. 39 (10): 487–95. doi:10.1016/j.tibs.2014.08.006. PMID 25239056.
- ↑ Kedersha N, Anderson P (November 2002). "Stress granules: sites of mRNA triage that regulate mRNA stability and translatability". Biochemical Society Transactions. 30 (Pt 6): 963–9. doi:10.1042/bst0300963. PMID 12440955.
- 1 2 An S, Kumar R, Sheets ED, Benkovic SJ (April 2008). "Reversible compartmentalization of de novo purine biosynthetic complexes in living cells". Science. 320 (5872): 103–6. Bibcode:2008Sci...320..103A. doi:10.1126/science.1152241. PMID 18388293. S2CID 24119538.
- ↑ Walter H, Brooks DE (March 1995). "Phase separation in cytoplasm, due to macromolecular crowding, is the basis for microcompartmentation". FEBS Letters. 361 (2–3): 135–9. doi:10.1016/0014-5793(95)00159-7. PMID 7698310. S2CID 8843457.
- ↑ Walter H, Brooks D, Srere P, eds. (October 1999). Microcompartmentation and Phase Separation in Cytoplasm. Vol. 192 (1 ed.). Academic Press.
- ↑ Brooks DE (1999). "Can Cytoplasm Exist without Undergoing Phase Separation?". Microcompartmentation and Phase Separation in Cytoplasm. International Review of Cytology. Vol. 192. pp. 321–330. doi:10.1016/S0074-7696(08)60532-X. ISBN 9780123645968. ISSN 0074-7696. PMID 10610362.
- ↑ Walter, Harry (1999). "Consequences of Phase Separation in Cytoplasm". Microcompartmentation and Phase Separation in Cytoplasm. International Review of Cytology. Vol. 192. pp. 331–343. doi:10.1016/S0074-7696(08)60533-1. ISBN 9780123645968. ISSN 0074-7696. PMID 10610363.
- ↑ Sear, Richard P. (1999). "Phase behavior of a simple model of globular proteins". The Journal of Chemical Physics. 111 (10): 4800–4806. arXiv:cond-mat/9904426. Bibcode:1999JChPh.111.4800S. doi:10.1063/1.479243. ISSN 0021-9606. S2CID 15005765.
- 1 2 Stradner A, Sedgwick H, Cardinaux F, Poon WC, Egelhaaf SU, Schurtenberger P (November 2004). "Equilibrium cluster formation in concentrated protein solutions and colloids" (PDF). Nature. 432 (7016): 492–5. Bibcode:2004Natur.432..492S. doi:10.1038/nature03109. PMID 15565151. S2CID 4373710.
- ↑ Iborra FJ (April 2007). "Can visco-elastic phase separation, macromolecular crowding and colloidal physics explain nuclear organisation?". Theoretical Biology & Medical Modelling. 4 (15): 15. doi:10.1186/1742-4682-4-15. PMC 1853075. PMID 17430588.
- 1 2 3 Sear RP (May 2007). "Dishevelled: a protein that functions in living cells by phase separating". Soft Matter. 3 (6): 680–684. Bibcode:2007SMat....3..680S. doi:10.1039/b618126k. PMID 32900127.
- ↑ Sear RP (2008). "Phase separation of equilibrium polymers of proteins in living cells". Faraday Discussions. 139: 21–34, discussion 105–28, 419–20. Bibcode:2008FaDi..139...21S. doi:10.1039/b713076g. PMID 19048988.
- ↑ Dumetz AC, Chockla AM, Kaler EW, Lenhoff AM (January 2008). "Protein phase behavior in aqueous solutions: crystallization, liquid–liquid phase separation, gels, and aggregates". Biophysical Journal. 94 (2): 570–83. Bibcode:2008BpJ....94..570D. doi:10.1529/biophysj.107.116152. PMC 2157236. PMID 18160663.
- 1 2 3 4 Brangwynne CP, Eckmann CR, Courson DS, Rybarska A, Hoege C, Gharakhani J, et al. (June 2009). "Germline P granules are liquid droplets that localize by controlled dissolution/condensation". Science. 324 (5935): 1729–32. Bibcode:2009Sci...324.1729B. doi:10.1126/science.1172046. PMID 19460965. S2CID 42229928.
- ↑ Larson AG, Elnatan D, Keenen MM, Trnka MJ, Johnston JB, Burlingame AL, et al. (July 2017). "Liquid droplet formation by HP1α suggests a role for phase separation in heterochromatin". Nature. 547 (7662): 236–240. Bibcode:2017Natur.547..236L. doi:10.1038/nature22822. PMC 5606208. PMID 28636604.
- ↑ Nott TJ, Petsalaki E, Farber P, Jervis D, Fussner E, Plochowietz A, et al. (March 2015). "Phase transition of a disordered nuage protein generates environmentally responsive membraneless organelles". Molecular Cell. 57 (5): 936–947. doi:10.1016/j.molcel.2015.01.013. PMC 4352761. PMID 25747659.
- ↑ Patel A, Lee HO, Jawerth L, Maharana S, Jahnel M, Hein MY, et al. (August 2015). "A Liquid-to-Solid Phase Transition of the ALS Protein FUS Accelerated by Disease Mutation". Cell. 162 (5): 1066–77. doi:10.1016/j.cell.2015.07.047. PMID 26317470.
- 1 2 Feric M, Vaidya N, Harmon TS, Mitrea DM, Zhu L, Richardson TM, et al. (June 2016). "Coexisting Liquid Phases Underlie Nucleolar Subcompartments". Cell. 165 (7): 1686–1697. doi:10.1016/j.cell.2016.04.047. PMC 5127388. PMID 27212236.
- ↑ Riback JA, Zhu L, Ferrolino MC, Tolbert M, Mitrea DM, Sanders DW, et al. (2019-10-22). "Composition dependent phase separation underlies directional flux through the nucleolus". bioRxiv: 809210. doi:10.1101/809210.
- 1 2 Li P, Banjade S, Cheng HC, Kim S, Chen B, Guo L, et al. (March 2012). "Phase transitions in the assembly of multivalent signalling proteins". Nature. 483 (7389): 336–40. Bibcode:2012Natur.483..336L. doi:10.1038/nature10879. PMC 3343696. PMID 22398450.
- 1 2 3 Banani SF, Lee HO, Hyman AA, Rosen MK (May 2017). "Biomolecular condensates: organizers of cellular biochemistry". Nature Reviews. Molecular Cell Biology. 18 (5): 285–298. doi:10.1038/nrm.2017.7. PMC 7434221. PMID 28225081. S2CID 37694361.
- ↑ Wheeler RJ, Hyman AA (May 2018). "Controlling compartmentalization by non-membrane-bound organelles". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 373 (1747): 4666–4684. doi:10.1098/rstb.2017.0193. PMC 5904305. PMID 29632271.
- ↑ Garaizar A, Sanchez-Burgos I, Collepardo-Guevara R, Espinosa JR (October 2020). "Expansion of Intrinsically Disordered Proteins Increases the Range of Stability of Liquid–Liquid Phase Separation". Molecules. 25 (20): 4705. doi:10.3390/molecules25204705. PMC 7587599. PMID 33076213.
- ↑ Kato M, McKnight SL (March 2017). "Cross-β Polymerization of Low Complexity Sequence Domains". Cold Spring Harbor Perspectives in Biology. 9 (3): a023598. doi:10.1101/cshperspect.a023598. PMC 5334260. PMID 27836835.
- ↑ Bienz M (August 2020). "Head-to-Tail Polymerization in the Assembly of Biomolecular Condensates". Cell. 182 (4): 799–811. doi:10.1016/j.cell.2020.07.037. PMID 32822572. S2CID 221198567.
- ↑ Nakano A, Trie R, Tateishi K (January 1997). "Glycogen-Surfactant Complexes: Phase Behavior in a Water/Phytoglycogen/Sodium Dodecyl Sulfate (SDS) System". Bioscience, Biotechnology, and Biochemistry. 61 (12): 2063–8. doi:10.1271/bbb.61.2063. PMID 27396883.
- ↑ Esposito, Mark; Fang, Cao; Cook, Katelyn C.; Park, Nana; Wei, Yong; Spadazzi, Chiara; Bracha, Dan; Gunaratna, Ramesh T.; Laevsky, Gary; DeCoste, Christina J.; Slabodkin, Hannah (March 2021). "TGF-β-induced DACT1 biomolecular condensates repress Wnt signalling to promote bone metastasis". Nature Cell Biology. 23 (3): 257–267. doi:10.1038/s41556-021-00641-w. ISSN 1476-4679. PMC 7970447. PMID 33723425.
- 1 2 Schaefer KN, Peifer M (February 2019). "Wnt/Beta-Catenin Signaling Regulation and a Role for Biomolecular Condensates". Developmental Cell. 48 (4): 429–444. doi:10.1016/j.devcel.2019.01.025. PMC 6386181. PMID 30782412.
- 1 2 Gammons M, Bienz M (April 2018). "Multiprotein complexes governing Wnt signal transduction". Current Opinion in Cell Biology. 51 (1): 42–49. doi:10.1016/j.ceb.2017.10.008. PMID 29153704.
- ↑ Muthunayake NS, Tomares DT, Childers WS, Schrader JM (November 2020). "Phase-separated bacterial ribonucleoprotein bodies organize mRNA decay". Wiley Interdisciplinary Reviews. RNA. 11 (6): e1599. doi:10.1002/wrna.1599. PMC 7554086. PMID 32445438.
- ↑ Dorone, Yanniv; Boeynaems, Steven; Jin, Benjamin; Bossi, Flavia; Flores, Eduardo; Lazarus, Elena; Michiels, Emiel; De Decker, Mathias; Baatsen, Pieter; Holehouse, Alex S.; Sukenik, Shahar; Gitler, Aaron D.; Rhee, Seung Y. (July 2021). "A prion-like protein regulator of seed germination undergoes hydration-dependent phase separation". Cell. 184 (16): 4284–4298.e27. doi:10.1016/j.cell.2021.06.009. PMC 8513799. PMID 34233164. S2CID 221096771.
- ↑ Case LB, Ditlev JA, Rosen MK (May 2019). "Regulation of Transmembrane Signaling by Phase Separation". Annual Review of Biophysics. 48 (1): 465–494. doi:10.1146/annurev-biophys-052118-115534. PMC 6771929. PMID 30951647.
- ↑ Muschol, Martin; Rosenberger, Franz (1997). "Liquid–liquid phase separation in supersaturated lysozyme solutions and associated precipitate formation/crystallization". The Journal of Chemical Physics. 107 (6): 1953–1962. Bibcode:1997JChPh.107.1953M. doi:10.1063/1.474547. ISSN 0021-9606.
- ↑ Patterson M, Vogel HJ, Prenner EJ (February 2016). "Biophysical characterization of monofilm model systems composed of selected tear film phospholipids". Biochimica et Biophysica Acta (BBA) - Biomembranes. 1858 (2): 403–14. doi:10.1016/j.bbamem.2015.11.025. PMID 26657693.
- 1 2 Tang L (February 2019). "Optogenetic tools light up phase separation". Nature Methods (Paper). 16 (2): 139. doi:10.1038/s41592-019-0310-5. PMID 30700901. S2CID 59525729.(subscription required)
- ↑ Hyman AA, Weber CA, Jülicher F (2014-10-11). "Liquid–liquid phase separation in biology". Annual Review of Cell and Developmental Biology. 30 (1): 39–58. doi:10.1146/annurev-cellbio-100913-013325. PMID 25288112.
- ↑ McSwiggen DT, Mir M, Darzacq X, Tjian R (December 2019). "Evaluating phase separation in live cells: diagnosis, caveats, and functional consequences". Genes & Development. 33 (23–24): 1619–1634. doi:10.1101/gad.331520.119. PMC 6942051. PMID 31594803.
- ↑ Posey AE, Holehouse AS, Pappu RV (2018). "Phase Separation of Intrinsically Disordered Proteins". Intrinsically Disordered Proteins. Methods in Enzymology. Vol. 611. Elsevier. pp. 1–30. doi:10.1016/bs.mie.2018.09.035. ISBN 978-0-12-815649-0. PMID 30471685.
- ↑ Woodruff JB, Hyman AA, Boke E (February 2018). "Organization and Function of Non-dynamic Biomolecular Condensates". Trends in Biochemical Sciences. 43 (2): 81–94. doi:10.1016/j.tibs.2017.11.005. PMID 29258725.
- ↑ Boeynaems S, Alberti S, Fawzi NL, Mittag T, Polymenidou M, Rousseau F, et al. (June 2018). "Protein Phase Separation: A New Phase in Cell Biology". Trends in Cell Biology. 28 (6): 420–435. doi:10.1016/j.tcb.2018.02.004. PMC 6034118. PMID 29602697.
- ↑ de Swaan Arons, J.; Diepen, G. A. M. (2010). "Immiscibility of gases. The system He-Xe: (Short communication)". Recueil des Travaux Chimiques des Pays-Bas. 82 (8): 806. doi:10.1002/recl.19630820810. ISSN 0165-0513.
- ↑ de Swaan Arons, J.; Diepen, G. A. M. (1966). "Gas—Gas Equilibria". J. Chem. Phys. 44 (6): 2322. Bibcode:1966JChPh..44.2322D. doi:10.1063/1.1727043.
- ↑ Bayro MJ, Daviso E, Belenky M, Griffin RG, Herzfeld J (January 2012). "An amyloid organelle, solid-state NMR evidence for cross-β assembly of gas vesicles". The Journal of Biological Chemistry. 287 (5): 3479–84. doi:10.1074/jbc.M111.313049. PMC 3271001. PMID 22147705.
- ↑ Elbaum-Garfinkle S, Kim Y, Szczepaniak K, Chen CC, Eckmann CR, Myong S, Brangwynne CP (June 2015). "The disordered P granule protein LAF-1 drives phase separation into droplets with tunable viscosity and dynamics". Proceedings of the National Academy of Sciences of the United States of America. 112 (23): 7189–94. Bibcode:2015PNAS..112.7189E. doi:10.1073/pnas.1504822112. PMC 4466716. PMID 26015579.
- ↑ Heidenreich M, Georgeson JM, Locatelli E, Rovigatti L, Nandi SK, Steinberg A, et al. (September 2020). "Designer protein assemblies with tunable phase diagrams in living cells". Nature Chemical Biology. 16 (9): 939–945. doi:10.1038/s41589-020-0576-z. hdl:11573/1435875. PMID 32661377. S2CID 220507058.
- ↑ Aguzzi A, Altmeyer M (July 2016). "Phase Separation: Linking Cellular Compartmentalization to Disease". Trends in Cell Biology. 26 (7): 547–558. doi:10.1016/j.tcb.2016.03.004. PMID 27051975.
- ↑ Shin Y, Brangwynne CP (September 2017). "Liquid phase condensation in cell physiology and disease". Science. 357 (6357): eaaf4382. doi:10.1126/science.aaf4382. PMID 28935776. S2CID 3693853.
- ↑ Alberti S, Hyman AA (October 2016). "Are aberrant phase transitions a driver of cellular aging?". BioEssays. 38 (10): 959–68. doi:10.1002/bies.201600042. PMC 5108435. PMID 27554449.
- ↑ P. Ivanov, N. Kedersha, and P. Anderson, “Stress granules and processing bodies in translational control,” Cold Spring Harbor Perspectives in Biology, vol. 11, no. 5, 2019.
- ↑ C. P. Brangwynne, T. J. Mitchison, and A. A. Hyman, “Active liquid-like behavior of nucleoli determines their size and shape in Xenopus laevis oocytes,” Proceedings of the National Academy of Sciences of the United States of America, vol. 108, no. 11, pp. 4334–4339, 2011.
- ↑ D. Bracha, M. T. Walls, and C. P. Brangwynne, “Probing and engineering liquid-phase organelles,” Nature Biotechnology, vol. 37, no. 12, pp. 1435–1445, 2019.
- ↑ Choi, J.M.; Dar, F.; Pappu, R.V. (2019). "LASSI: A lattice model for simulating phase transitions of multivalent proteins". PLOS Computational Biology. 15 (10): e1007028. Bibcode:2019PLSCB..15E7028C. doi:10.1371/journal.pcbi.1007028. PMC 6822780. PMID 31634364.
- ↑ Hastings, R.L.; Boeynaems, S. (June 2021). "Designer Condensates: A Toolkit for the Biomolecular Architect". Journal of Molecular Biology. 433 (12): 166837. doi:10.1016/j.jmb.2021.166837. PMID 33539874. S2CID 231819801.
- ↑ Tejedor, R.; Garaizar, A.; Ramı, J. (December 2021). "RNA modulation of transport properties and stability in phase-separated condensates". Biophysical Journal. 120 (23): 5169–5186. Bibcode:2021BpJ...120.5169T. doi:10.1016/j.bpj.2021.11.003. PMC 8715277. PMID 34762868.
- ↑ C. D. Reinkemeier and E. A. Lemke, “Synthetic biomolecular condensates to engineer eukaryotic cells,” Current Opinion in Chemical Biology, vol. 64, pp. 174–181, 2021.
- ↑ D. Bracha, M. T. Walls, M. T. Wei, L. Zhu, M. Kurian, J. L. Avalos, J. E. Toettcher, and C. P. Brangwynne, “Mapping Local and Global Liquid Phase Behavior in Living Cells Using Photo-Oligomerizable Seeds,” Cell, vol. 175, no. 6, pp. 1467–1480.e13, 2018.
- ↑ H. Zhang, C. Aonbangkhen, E. V. Tarasovetc, E. R. Ballister, D. M.Chenoweth, and M. A. Lampson, “Optogenetic control of kinetochore function,” Nature Chemical Biology, vol. 13, pp. 1096–1101, Aug 2017.
- 1 2 Y. Shin, Y. C. Chang, D. S. Lee, J. Berry, D. W. Sanders, P. Ronceray, N. S.Wingreen, M. Haataja, and C. P. Brangwynne, “Liquid Nuclear Condensates Mechanically Sense and Restructure the Genome,” Cell, vol. 175, no. 6,pp. 1481–1491.e13, 2018.
- 1 2 M. Yoshikawa and S. Tsukiji, “Modularly Built Synthetic Membraneless Organelles Enabling Targeted Protein Sequestration and Release,” Biochemistry, Oct 2021.
- ↑ Y. Shin and C. P. Brangwynne, “Liquid phase condensation in cell physiology and disease,” Science, vol. 357, Sep 2017.
- ↑ Methods to Study Phase-Separated Condensates and the Underlying Molecular Interactions
- ↑ Maziar Heidari et al., A topology framework for macromolecular complexes and condensates. Nano Research (2022)
- 1 2 3 Saar, Kadi L.; Qian, Daoyuan; Good, Lydia L.; Morgunov, Alexey S.; Collepardo-Guevara, Rosana; Best, Robert B.; Knowles, Tuomas P. J. (12 May 2023). "Theoretical and Data-Driven Approaches for Biomolecular Condensates". Chemical Reviews. 123 (14): 8988–9009. doi:10.1021/acs.chemrev.2c00586. eISSN 1520-6890. ISSN 0009-2665. PMC 10375482. PMID 37171907.
- ↑ Paloni, Matteo; Bailly, Rémy; Ciandrini, Luca; Barducci, Alessandro (16 September 2020). "Unraveling Molecular Interactions in Liquid–Liquid Phase Separation of Disordered Proteins by Atomistic Simulations". The Journal of Physical Chemistry B. 124 (41): 9009–9016. doi:10.1021/acs.jpcb.0c06288. eISSN 1520-5207. ISSN 1520-6106. PMID 32936641.
- ↑ Benayad, Zakarya; von Bülow, Sören; Stelzl, Lukas S.; Hummer, Gerhard (14 December 2020). "Simulation of FUS Protein Condensates with an Adapted Coarse-Grained Model". Journal of Chemical Theory and Computation. 17 (1): 525–537. doi:10.1021/acs.jctc.0c01064. eISSN 1549-9626. ISSN 1549-9618. PMC 7872324. PMID 33307683.
- ↑ Espinosa, Jorge R.; Joseph, Jerelle A.; Sanchez-Burgos, Ignacio; Garaizar, Adiran; Frenkel, Daan; Collepardo-Guevara, Rosana (June 2020). "Liquid network connectivity regulates the stability and composition of biomolecular condensates with many components". Proceedings of the National Academy of Sciences. 117 (24): 13238–13247. Bibcode:2020PNAS..11713238E. doi:10.1073/pnas.1917569117. eISSN 1091-6490. ISSN 0027-8424. PMC 7306995. PMID 32482873.
- ↑ Dignon, Gregory L.; Zheng, Wenwei; Kim, Young C.; Best, Robert B.; Mittal, Jeetain (24 January 2018). "Sequence determinants of protein phase behavior from a coarse-grained model". PLOS Computational Biology. 14 (1): e1005941. Bibcode:2018PLSCB..14E5941D. doi:10.1371/journal.pcbi.1005941. eISSN 1553-7358. PMC 5798848. PMID 29364893.
- ↑ Vernon, Robert McCoy; Chong, Paul Andrew; Tsang, Brian; Kim, Tae Hun; Bah, Alaji; Farber, Patrick; Lin, Hong; Forman-Kay, Julie Deborah (9 February 2018). "Pi-Pi contacts are an overlooked protein feature relevant to phase separation". eLife. 7. doi:10.7554/eLife.31486. eISSN 2050-084X. PMC 5847340. PMID 29424691.
- 1 2 3 4 Joseph, Jerelle A.; Reinhardt, Aleks; Aguirre, Anne; Chew, Pin Yu; Russell, Kieran O.; Espinosa, Jorge R.; Garaizar, Adiran; Collepardo-Guevara, Rosana (22 November 2021). "Physics-driven coarse-grained model for biomolecular phase separation with near-quantitative accuracy". Nature Computational Science. 1 (11): 732–743. doi:10.1038/s43588-021-00155-3. eISSN 2662-8457. PMC 7612994. PMID 35795820.
- 1 2 Regy, Roshan Mammen; Thompson, Jacob; Kim, Young C.; Mittal, Jeetain (24 May 2021). "Improved coarse‐grained model for studying sequence dependent phase separation of disordered proteins". Protein Science. 30 (7): 1371–1379. doi:10.1002/pro.4094. eISSN 1469-896X. ISSN 0961-8368. PMC 8197430. PMID 33934416.
- 1 2 Dannenhoffer-Lafage, Thomas; Best, Robert B. (20 April 2021). "A Data-Driven Hydrophobicity Scale for Predicting Liquid–Liquid Phase Separation of Proteins". The Journal of Physical Chemistry B. 125 (16): 4046–4056. doi:10.1021/acs.jpcb.0c11479. eISSN 1520-5207. ISSN 1520-6106. PMID 33876938. S2CID 233309675.
- 1 2 Latham, Andrew P.; Zhang, Bin (7 April 2021). "Consistent Force Field Captures Homologue-Resolved HP1 Phase Separation". Journal of Chemical Theory and Computation. 17 (5): 3134–3144. doi:10.1021/acs.jctc.0c01220. eISSN 1549-9626. ISSN 1549-9618. PMC 8119372. PMID 33826337.
- ↑ Tesei, Giulio; Schulze, Thea K.; Crehuet, Ramon; Lindorff-Larsen, Kresten (29 October 2021). "Accurate model of liquid–liquid phase behavior of intrinsically disordered proteins from optimization of single-chain properties". Proceedings of the National Academy of Sciences. 118 (44). Bibcode:2021PNAS..11811696T. doi:10.1073/pnas.2111696118. eISSN 1091-6490. ISSN 0027-8424. PMC 8612223. PMID 34716273.
- 1 2 Farag, Mina; Cohen, Samuel R.; Borcherds, Wade M.; Bremer, Anne; Mittag, Tanja; Pappu, Rohit V. (13 December 2022). "Condensates formed by prion-like low-complexity domains have small-world network structures and interfaces defined by expanded conformations". Nature Communications. 13 (1): 7722. Bibcode:2022NatCo..13.7722F. doi:10.1038/s41467-022-35370-7. eISSN 2041-1723. PMC 9748015. PMID 36513655.
- 1 2 3 Valdes-Garcia, Gilberto; Heo, Lim; Lapidus, Lisa J.; Feig, Michael (6 January 2023). "Modeling Concentration-dependent Phase Separation Processes Involving Peptides and RNA via Residue-Based Coarse-Graining". Journal of Chemical Theory and Computation. 19 (2): 669–678. doi:10.1021/acs.jctc.2c00856. eISSN 1549-9626. ISSN 1549-9618. PMC 10323037. PMID 36607820.
- ↑ Tesei, Giulio; Lindorff-Larsen, Kresten (17 January 2023). "Improved predictions of phase behaviour of intrinsically disordered proteins by tuning the interaction range". Open Research Europe. 2: 94. doi:10.12688/openreseurope.14967.2. eISSN 2732-5121. PMC 10450847. PMID 37645312.
- ↑ Mittag, Tanja; Pappu, Rohit V. (June 2022). "A conceptual framework for understanding phase separation and addressing open questions and challenges". Molecular Cell. 82 (12): 2201–2214. doi:10.1016/j.molcel.2022.05.018. ISSN 1097-2765. PMC 9233049. PMID 35675815. S2CID 249488875.
- ↑ Borcherds, Wade; Bremer, Anne; Borgia, Madeleine B; Mittag, Tanja (April 2021). "How do intrinsically disordered protein regions encode a driving force for liquid–liquid phase separation?". Current Opinion in Structural Biology. 67: 41–50. doi:10.1016/j.sbi.2020.09.004. ISSN 0959-440X. PMC 8044266. PMID 33069007.
- ↑ Ghosh, Soumyadeep (April 28, 2023). "Scaffolds and Clients". CD-CODE Encyclopedia. Retrieved May 28, 2023.
- ↑ Regy, Roshan Mammen; Dignon, Gregory L; Zheng, Wenwei; Kim, Young C; Mittal, Jeetain (2 December 2020). "Sequence dependent phase separation of protein-polynucleotide mixtures elucidated using molecular simulations". Nucleic Acids Research. 48 (22): 12593–12603. doi:10.1093/nar/gkaa1099. eISSN 1362-4962. ISSN 0305-1048. PMC 7736803. PMID 33264400.
- ↑ Lebold, Kathryn M.; Best, Robert B. (23 March 2022). "Tuning Formation of Protein–DNA Coacervates by Sequence and Environment". The Journal of Physical Chemistry B. 126 (12): 2407–2419. doi:10.1021/acs.jpcb.2c00424. eISSN 1520-5207. ISSN 1520-6106. PMID 35317553.
- ↑ Leicher, Rachel; Osunsade, Adewola; Chua, Gabriella N. L.; Faulkner, Sarah C.; Latham, Andrew P.; Watters, John W.; Nguyen, Tuan; Beckwitt, Emily C.; Christodoulou-Rubalcava, Sophia; Young, Paul G.; Zhang, Bin; David, Yael; Liu, Shixin (28 April 2022). "Single-stranded nucleic acid binding and coacervation by linker histone H1". Nature Structural & Molecular Biology. 29 (5): 463–471. doi:10.1038/s41594-022-00760-4. eISSN 1545-9985. ISSN 1545-9993. PMC 9117509. PMID 35484234.
- ↑ Farr, Stephen E.; Woods, Esmae J.; Joseph, Jerelle A.; Garaizar, Adiran; Collepardo-Guevara, Rosana (17 May 2021). "Nucleosome plasticity is a critical element of chromatin liquid–liquid phase separation and multivalent nucleosome interactions". Nature Communications. 12 (1): 2883. Bibcode:2021NatCo..12.2883F. doi:10.1038/s41467-021-23090-3. eISSN 2041-1723. PMC 8129070. PMID 34001913.
- ↑ Latham, Andrew P.; Zhang, Bin (February 2022). "Unifying coarse-grained force fields for folded and disordered proteins". Current Opinion in Structural Biology. 72: 63–70. doi:10.1016/j.sbi.2021.08.006. ISSN 0959-440X. PMC 9057422. PMID 34536913.
Further reading
- Ditlev JA, Case LB, Rosen MK (November 2018). "Who's In and Who's Out-Compositional Control of Biomolecular Condensates". Journal of Molecular Biology. 430 (23): 4666–4684. doi:10.1016/j.jmb.2018.08.003. PMC 6204295. PMID 30099028.
- Banani SF, Lee HO, Hyman AA, Rosen MK (May 2017). "Biomolecular condensates: organizers of cellular biochemistry". Nature Reviews. Molecular Cell Biology. 18 (5): 285–298. doi:10.1038/nrm.2017.7. PMC 7434221. PMID 28225081. S2CID 37694361.
- Hyman AA, Weber CA, Jülicher F (2014). "Liquid–liquid phase separation in biology". Annual Review of Cell and Developmental Biology. 30: 39–58. doi:10.1146/annurev-cellbio-100913-013325. PMID 25288112.
- Dolgin E (March 2018). "What lava lamps and vinaigrette can teach us about cell biology". Nature. 555 (7696): 300–302. Bibcode:2018Natur.555..300D. doi:10.1038/d41586-018-03070-2. PMID 29542707.