Mal'ta–Buret' culture ivory figurines (c. 24,000 BP-c. 15,000 BP). Some of the figurines wear hooded overalls with decorative stripes.[1]
[2] |
.jpg.webp)
.png.webp)

In archaeogenetics, the term Ancient North Eurasian (ANE) is the name given to an ancestral component that represents the lineage of the people of the Mal'ta–Buret' culture (c. 24,000 BP) and populations closely related to them, such as the Upper Paleolithic individuals from Afontova Gora in Siberia.[6][7] Genetic studies indicate that the ANE are closely related to the Ancient North Siberians (ANS) represented by two ancient specimens from the preceding Yana Culture (c. 32,000 BP). The ANE can either be considered to descend from the earlier ANS population, or that both ANE and ANS are closely related, albeit differentiated, sister lineages, with both having originated from an 'Early West Eurasian' hunter-gatherer lineage (represented by Kostenki-14, c. 40,000 BP),[8] which absorbed an 'Early East Eurasian' population (represented by the Tianyuan man, c. 40,000 BP). The ANS and ANE each derive between 16% to 35% of their ancestry from an Early East Eurasian lineage and between 65% to 84% from an Early West Eurasian lineage.[lower-alpha 1][lower-alpha 2][9][10][11][12][13][14]
Around 20,000 to 25,000 years ago, a branch of Ancient North Eurasian people mixed with Ancient East Asians, which led to the emergence of Ancestral Native American, Ancient Beringian and Ancient Paleo-Siberian populations. It is unknown exactly where this population admixture took place, and two opposing theories have put forth different migratory scenarios that united the Ancient North Eurasians with ancient East Asian populations.[15]
ANE ancestry has spread throughout Eurasia and the Americas in various migrations since the Upper Paleolithic, and more than half of the world's population today derives between 5 and 42% of their genomes from the Ancient North Eurasians.[16] Significant ANE ancestry can be found in Native Americans, as well as in regions of northern Europe, South Asia, Central Asia, and Siberia. It has been suggested that their mythology may have featured narratives shared by both Indo-European and some Native American cultures, such as the existence of a metaphysical world tree and a fable in which a dog guards the path to the afterlife.[17]
Genetic studies
Definition
The ANE lineage is defined by association with the "Mal'ta boy" (MA-1), the remains of an individual who lived during the Last Glacial Maximum, 24,000 years ago in central Siberia, discovered in the 1920s. Together with the Yana Rhinoceros Horn Site samples, and Afontova Gora individuals, they are collectively referred to as 'Ancient North Siberians'.[18][19]
_with_tomb_artifacts%252C_Hermitage_Museum%252C_Saint-Petersburg.jpg.webp)
The Ancient North Eurasians represent a distinct cluster of genetic diversity within the larger Eurasian gene pool, which is deeply related to Paleolithic and Mesolithic European hunter-gatherers.[22] It is suggested that the ANE ancestry found among modern human populations was largely contributed from a population linked to Afontova Gora (AG-3), rather than Malta (MA-1) or Yana.[23]
Formation
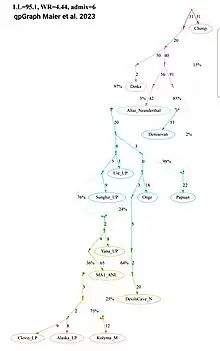
The formation of the Ancient North Eurasian/Siberian gene pool likely occurred very early via an 'Early West Eurasian' Upper Paleolithic (UP) lineage, deeply related to 'European hunter-gatherers', migrating along the "Northern route" into Siberia,[25] and subsequently merging with an 'Early East Eurasian' Initial Upper Paleolithic (IUP) lineage, basal to contemporary East and Southeast Asian populations. The ANE/ANS-associated samples from the Yana Rhinoceros Horn Site (31,600 BP) in Northeastern Siberia, the Mal'ta–Buret' culture, and the individuals associated with the Afontova Gora site, can be modeled to derive between c. 65% to 84% of their ancestry from Early West Eurasians (c. 38kya), with varying but significant contributions (c. 16% to 35%) from Early East Eurasians (>40kya). The exact percentage of each component vary depending on the used model. Their West Eurasian component can be associated with ancestry found among Paleolithic European hunter-gatherers, such as Kostenki-14, Sunghir, or the Peștera Muierii woman,[26] their East Eurasian component can be associated with ancestry found among a population related to the Paleolithic Tianyuan man, who is basal to contemporary East/Southeast Asians.[27][28][29][30][31][32][lower-alpha 3][lower-alpha 4][33][13][14]
Lipson and Reich (2017) modeled the Malta sample to derive ancestry from a West Eurasian source, with additional admixture from a lineage basal to East Asians (represented by Amis), while also noting the possibility for a reversed geneflow from Malta into East Asians, which however had less support with the available data.[33] Yang et al. 2020a,b, corroborated that both the Malta and Yana specimens formed from the merger of a sister lineage of the 'European hunter-gatherer' Kostenki-14, and from a lineage contemporary to the 'Basal-East Asian' Tianyuan man, while finding no evidence for a reversed geneflow from ANE into Tianyuan or modern East Asians.[34][35] Other studies could reproduce a shared affinity between ANE and the Tianyuan man (or other Upper Paleolithic East/Southeast Asian specimens), and also confirmed higher affinity between the Tianyuan man and Eastern European Hunter-Gatherers (EHG), which received significant amounts of ANE ancestry.[27] The 'Basal East Asian' (Tianyuan-like) ancestry among EHGs (Sidelkino) has been estimated to be around 12,9%.[36]
Grebenyuk et al. summarized that the Ancient North Eurasians descended from the 'Ancient North Siberian' Yana population, which were "Early Upper Paleolithic tribes of hunters" and linked to similar groups associated with contemporaneous Southern Siberian sites. These communities of Southern Siberian and Central Asian hunters belonged to one of the earliest migration waves of the anatomically modern humans into Siberia. The authors summarized that "the initial peopling of Northeastern Asia by the anatomically modern humans could have happened both from West to East and from South to North".[37] Sikora et al. notes that the Ancient North Eurasians (Malta and Afontova Gora individuals) are unlikely to be direct descendants of the 'Ancient North Siberian' Yana population; rather, the study argues, both are sister lineages sharing a common ancestor. According to Sikora et al., the Malta sample may additionally also have received some 'early Caucasus hunter-gatherer' geneflow (c. 11%).[19] This scenario is questioned by Maier et al. 2023, who state that this conclusion is contradicted by other published articles, and that the direction of gene flow as well as observed affinity between ANE and CHG populations cannot be demonstrated by analysis of admixture graphs, but need further investigation.[38]
Distribution


By c. 32kya, populations carrying ANE-related ancestry were probably widely distributed across northeast Eurasia. They may have expanded as far as Alaska and the Yukon, but were forced to abandon high latitude regions following the onset of harsher climatic conditions that came with the Last Glacial Maximum.[40]
Populations genetically similar to MA-1 and Afontova Gora were an important genetic contributor to Native Americans, Europeans, Ancient Central Asians, South Asians, and some East Asian groups, in order of significance.[41] Lazaridis et al. (2016:10) note "a cline of ANE ancestry across the east-west extent of Eurasia". A 2016 study found that the global maximum of ANE ancestry occurs in modern-day Kets, Mansi, Native Americans, and Selkups.[6][41]

The ancient Bronze-age-steppe Yamnaya and Afanasevo cultures were found to have a significant ANE component at c. 50%.[43][44] According to Moreno-Mayar et al. 2018 between 14% and 38% of Native American ancestry may originate from gene flow from the Mal'ta–Buret' (ANE) population. This difference is caused by the penetration of posterior "Neo-Siberian" migrations into the Americas, with the lowest percentages of ANE ancestry found in Inuit and Alaskan Natives, as these groups are the result of migrations into the Americas roughly 5,000 years ago.[45] Estimates for ANE ancestry among first wave Native Americans show higher percentages,[46] such as 42% for those belonging to the Andean region in South America.[46] The other gene flow in Native Americans (the remainder of their ancestry) was of an East Asian-related origin, specifically diverged from other East Asians c. 30,000 years ago.[29] Gene sequencing of another south-central Siberian people (Afontova Gora-2) dating to approximately 17,000 years ago, revealed similar autosomal genetic signatures to that of Mal'ta boy-1, suggesting that the region was continuously occupied by humans throughout the Last Glacial Maximum.[29]
Genomic studies also indicate that the ANE component was brought to Western Europe by people related to the Yamnaya culture, long after the Paleolithic.[43][6] It is reported in modern-day Europeans (10%–20%).[43][6] Earlier ANE ancestry is found in European hunter-gatherer populations through Paleolithic interactions with Eastern European Hunter-Gatherers, which resulted in populations such as Scandinavian Hunter-Gatherers. [47] Western Hunter-Gatherers of the Villabruna cluster also carried the Y-haplogroup R1b, derived from the Ancient North Eurasian haplogroup R*, indicating "an early link between Europe and the western edge of the Steppe Belt of Eurasia."[48]
A deer tooth pendant impregnated with the genetic material of an ANE woman was found in the Denisova Cave, and dated to circa 24,700 years before present. She is closely related to Mal'ta and Afontova Gora specimens, found further east.[42]
An early Neolithic Central Asian specimen (Tutkaul1) from Tajikistan was found to be primarily derived from Ancient North Eurasians with some additional Neolithic Iranian-related inputs. The sample is closely related to Afontova Gora 3 (AG3) and Mal’ta 1, as well as to the West Siberian hunter-gatherers (Tyumen and Sosnoviy). While the sample also displays affinity for Eastern hunter-gatherers (EHGs), AG3 was found to be closer to EHGs than Tutkaul1, who instead may be a good proxy for ANE-related ancestry among ancient populations from the Iran and the Turan region.[49]
The Ancient Tianyuan Man and modern East/Southeast Asian populations were found to lack Upper Paleolithic Western Eurasian or ANE-related admixture, suggesting "resistance of those groups to the incoming UP population movements", or alternatively a subsequent reexpansion from a genetically East Asian-like population reservoir.[50]
Groups partially derived from the Ancient North Eurasians
Native American contribution
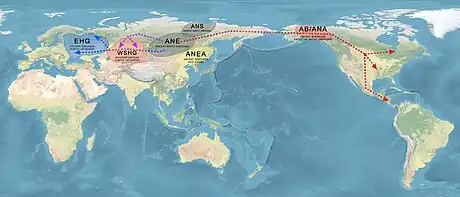

According to Jennifer Raff, the Ancient North Eurasian population mixed with a daughter population of ancient East Asians, who they encountered around 25,000 years ago, which lead to the emergence of Native American ancestral populations. However, the exact location where the admixture took place is unknown, and the migratory movements that united the two populations are a matter of debate.[15]
One theory supposes that Ancient North Eurasians migrated south to East Asia, or Southern Siberia, where they would have encountered and mixed with ancient East Asians. Genetic evidence from Lake Baikal in Mongolia supports this area as the location where the admixture took place.[51]
However, a third theory, the "Beringian standstill hypothesis", suggests that East Asians instead migrated north to Northeastern Siberia, where they mixed with ANE, and later diverged in Beringia, where distinct Native American lineages formed. This theory is supported by maternal and nuclear DNA evidence.[52] According to Grebenyuk, after 20,000 BP, a branch of Ancient East Asians migrated to Northeastern Siberia, and mixed with descendants of the ANE, leading to the emergence of Ancient Paleo-Siberian and Native American populations in Extreme Northeastern Asia.[53][37] However, the Beringian standstill hypothesis is not supported by paternal DNA evidence, which may reflect different population histories for paternal and maternal lineages in Native Americans, which is not uncommon and has been observed in other populations.[54]
The descendants of admixture between ANE and ancient East Asians include Ancient Beringian/Ancestral Native American, which are specific archaeogenetic lineages, based on the genome of an infant found at the Upward Sun River site (dubbed USR1), dated to 11,500 years ago.[55] The AB and the Ancestral Native American (ANA) lineage formed about 25,000 years ago, and subsequently diverged from each other, with the AB staying in the Beringian region, while the Ancestral Native Americans populated the Americas. The ANE genetic contribution to late-Paeolithic Ancestral Native Americans (USR1 specimen, dated to 11,500 BP in Alaska, and Clovis specimen, dated to 12,600 BP in Montana) is estimated at around 36.8%.[56] There are also the Ancient Paleo-Siberians, populations represented by the Late Upper Paeolithic Lake Baikal Ust'Kyakhta-3 (UKY) 14,050-13,770 BP. They carried 30% ANE ancestry and 70% East Asian ancestry.[57]
Jōmon people, the pre-Neolithic population of Japan, mainly derived their ancestry from East Asian lineages, but also received geneflow from the ANE-related "Ancient North Siberians" (represented by samples from the Yana Rhinoceros Horn Site) prior to the migration from the Asian mainland to the Japanese archipelago. Jōmon ancestry is still found among the inhabitants of present-day Japan: most markedly among the Ainu people, who are considered the direct descendants of the Jōmon people, and to a small, but significant degree among the majority of the Japanese population.[58][59]
Siberian and Asian Holocene populations
Altai hunter-gatherer is the name given to Middle Holocene Siberian hunter-gatherers within the Altai-Sayan region in Southern Siberia. They originated from the admixture of Paleo-Siberian and Ancient North Eurasian groups and show increased affinity towards Native Americans. Bronze Age groups from North and Inner Asia with significant ANE ancestry (e.g. Lake Baikal hunter-gatherers, Okunevo pastoralists) can be successfully modeled with Altai hunter-gatherers as a proximal ANE-derived ancestry source.[60] The Okunevos and Botai can be considered as direct descendents of the Ancient North Eurasians (ANE), specifically of the Malta-Buret people.[61]
West Siberian Hunter-Gatherer (WSHG) is a specific archaeogenetic lineage that was first reported by Narasimhan et al. (2019). It can be modeled as 20% EHG, 73% ANE and 6% Ancient Northeast Asian. Although only represented by three sampled hunter-gatherer individuals from Tyumen Oblast in the Russian Forest Zone east of the Urals dated ca. 5,000 BCE, high-levels of WSHG-like ancestry can be detected in various populations of Central Asia until the Bronze Age. The population of the Botai culture, while probably not directly descended from WSHG, displays a high affinity with the WSHG lineage.[62] The European-Siberian cline defined by Eastern hunter-gatherer-like ancestry streched from Central Europe to Siberia and was already established 10,000 years ago, including the West Siberian hunter-gatherers, all deriving their ancestry primarily from Paleolithic Siberians (ANE).[63]

Lake Baikal Holocene - Among the Ancient Northeast Asians (ANA) of the Neolithic to Early Bronze Age period, Baikal Eneolithic (Baikal_EN) and Baikal Early Bronze Age (Baikal_EBA) derived 6.4% to 20.1% ancestry from ANE, while the rest of their ancestry was derived from ANA. Fofonovo_EN near by Lake Baikal were mixture of 12-17% ANE ancestry and 83-87% ANA ancestry.[65]
Tarim mummies
A 2021 genetic study on the Tarim mummies found that they were primarily descended from a population represented by the Afontova Gora 3 specimen (AG3), genetically displaying "high affinity" with it.[66] The genetic profile of the Afontova Gora 3 individual represented about 72% of the ancestry of the Tarim mummies, while the remaining 28% of their ancestry was derived from a population represented by the Baikal EBA (Early Bronze Age Northeast Asian Baikal populations).[67]
The Tarim mummies are thus one of the rare Holocene populations who derive most of their ancestry from the Ancient North Eurasians (ANE, specifically the Mal'ta and Afontova Gora populations), despite their distance in time (around 14,000 years).[64] Having survived in a type of "genetic bottleneck" in the Tarim basin where they preserved and perpetuated their ANE ancestry, the Tarim mummies, more than any other ancient populations, can be considered as "the best representatives" of the Ancient North Eurasians.[64]
European populations
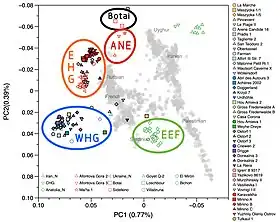
Lazaridis et al. (2014) detected ANE ancestry among modern European populations in proportions up to 20%.[68] In ancient European populations, the ANE genetic component is visible in tests of the Yamnaya people[43] but not of Western or Central Europeans predating the Corded Ware culture:[69] ANE ancestry was introduced in the European gene pool with the Eastern Hunter-Gatherer (EHG) lineage which derived significant ancestry from the ANE, c. 70%, with the remaining ancestry from a group more closely related to, but distinct from, Western Hunter-Gatherers (WHGs).[41][70][49] It is represented by multiple individuals, such as from Yuzhny Oleny in Karelia, one of Y-haplogroup R1a-M417, dated c. 8.4 kya, the other of Y-haplogroup J, dated c. 7.2 kya; and one individual from Samara, of Y-haplogroup R1b-P297, dated c. 7.6 kya, as well as individuals from Sidelkino and Popovo. After the end of the Last Glacial Maximum, the Western Hunter-Gatherers (WHG) and EHG lineages merged in Eastern Europe, accounting for early presence of ANE-derived ancestry in Mesolithic Europe. Evidence suggests that as Ancient North Eurasians migrated westward from Eastern Siberia, they absorbed Western Hunter-Gatherers and other West Eurasian populations as well.[71][49]
Scandinavian Hunter-Gatherer (SHG) is represented by several individuals buried at Motala, Sweden ca. 6000 BC. They were descended from Western Hunter-Gatherers who initially settled Scandinavia from the south, and received later admixture from EHG who entered Scandinavia from the north through the coast of Norway.[72][43][73][47][74]
Western Steppe Herders (WSH) is the name given to a distinct ancestral component that represents descent closely related to the Yamnaya culture of the Pontic–Caspian steppe.[lower-alpha 5] This ancestry is often referred to as Yamnaya ancestry or Steppe ancestry, and was formed from EHG and CHG (Caucasus hunter-gatherer) in about equal proportions.[28]
Phenotype prediction
.png.webp)
.jpg.webp)
Genomic studies by Raghavan et al. (2014) and Fu et al. (2016) suggested that Mal'ta boy may have had brown eyes, and relatively dark hair and dark skin,[76][77] while cautioning that this analysis was based on an extremely low coverage of DNA that might not give an accurate prediction of pigmentation.[78] Mathieson, et al. (2018) could not determine if Mal'ta 1 boy had the derived allele associated with blond hair in ANE descendants, as they could obtain no coverage for this SNP.[79]
Anthropologic research

Kozintsev (2020) argues that the historical Southern Siberian Okunevo population, which derives most of their ancestry from Ancient North Eurasians and their closest relatives, as possessing a distinct craniometric phenotype, which he dubbed "Americanoid", which represents the variation of the first humans in Siberia. He further argues that "As the geography and chronology of the ANE component show, it is misleading to describe it as Western Eurasian and associate it solely with ancient Caucasoids. To all appearances, it emerged before the Caucasoid-Mongoloid split."[44]
Zhang et al. (2021) proposed that the 'Western' like features of the earlier Tarim mummies could be attributed to their Ancient North Eurasian ancestry.[80] Previous craniometric analyses on the early Tarim mummies found that they formed their own cluster, and clustered with neither European-related Steppe pastoralists of the Andronovo and Afanasievo cultures, nor with inhabitants of the Western Asian BMAC culture, nor with East Asian populations further east.[81]
Evolution of blond hair
Blond hair is associated with a single nucleotide polymorphism, the mutated allele rs12821256 of the KITLG gene.[82][83][84][85][86] The earliest known individual with this allele is a female south-central Siberian ANE individual from the Afontova Gora 3 site, which is dated to c. 17,000 before present (the earlier ANE Mal'ta boy lacks the sequence coverage to make this determination).[87] The allele then appears later in ANE-derived Eastern Hunter-Gatherer (EHG) populations at Samara, Motala and Ukraine, circa 10,000 BP, and then in populations with Steppe ancestry.[79] Mathieson, et al. (2018) thus argued that this allele originated in the Ancient North Eurasian population, before spreading to western Eurasia.[79]
Geneticist David Reich said that the KITLG gene for blond hair probably entered continental Europe in a population migration wave from the Eurasian steppe, by a population carrying substantial Ancient North Eurasian ancestry.[88] Hanel and Carlberg (2020) likewise report that populations derived Ancient North Eurasian ancestry, specifically the Eastern Hunter-Gatherers and the Yamnayas, were responsible for transmitting this gene to Europeans.[89] The gene was also found among the Tarim mummies.[90]
.png.webp)
Comparative mythology


Since the term 'Ancient North Eurasian' refers to a genetic bridge of connected mating networks, scholars of comparative mythology have argued that they probably shared myths and beliefs that could be reconstructed via the comparison of stories attested within cultures that were not in contact for millennia and stretched from the Pontic–Caspian steppe to the American continent.[17]
The mytheme of the dog guarding the Otherworld possibly stems from an older Ancient North Eurasian belief, as suggested by similar motifs found in Indo-European, Native American and Siberian mythology. In Siouan, Algonquian, Iroquoian, and in Central and South American beliefs, a fierce guard dog was located in the Milky Way, perceived as the path of souls in the afterlife, and getting past it was a test. The Siberian Chukchi and Tungus believed in a guardian-of-the-afterlife dog and a spirit dog that would absorb the dead man's soul and act as a guide in the afterlife. In Indo-European myths, the figure of the dog is embodied by Cerberus, Sarvarā, and Garmr. In Zoroastrianism, two four-eyed dogs guard the bridge to the afterlife called Chinvat Bridge. Anthony and Brown note that it might be one of the oldest mythemes recoverable through comparative mythology.[17]
A second canid-related series of beliefs, myths and rituals connected dogs with healing rather than death. For instance, Ancient Near Eastern and Turkic-Kipchaq myths are prone to associate dogs with healing and generally categorised dogs as impure. A similar myth-pattern is assumed for the Eneolithic site of Botai in Kazakhstan, dated to 3500 BC, which might represent the dog as absorber of illness and guardian of the household against disease and evil. In Mesopotamia, the goddess Nintinugga, associated with healing, was accompanied or symbolized by dogs. Similar absorbent-puppy healing and sacrifice rituals were practiced in Greece and Italy, among the Hittites, again possibly influenced by Near Eastern traditions.[17]
See also
References
- 1 2 Bednarik, Robert G. (2013). "Pleistocene Palaeoart of Asia". Arts. 2 (2): 46–76. doi:10.3390/arts2020046.
- ↑ Lbova, Liudmila (2021). "The Siberian Palaeolithic Site of Mal'ta: A Unique Source for The Study of Childhood Archaeology". Evolutionary Human Sciences. 2: e9. doi:10.1017/ehs.2021.5. PMC 10427291. PMID 37588521. S2CID 231980510.
- ↑ "Paleolithic to Bronze Age Siberians Reveal Connections with First Americans and across Eurasia" (PDF). CELL. 2020.
- ↑ Yang, Melinda A. (6 January 2022). "A genetic history of migration, diversification, and admixture in Asia". Human Population Genetics and Genomics. 2 (1): 1–32. doi:10.47248/hpgg2202010001. ISSN 2770-5005.
- ↑ Willerslev, Eske; Meltzer, David J. (June 2021). "Willerslev, E., Meltzer, D.J. Peopling of the Americas as inferred from ancient genomics. Nature 594, 356–364 (2021)". Nature. 594 (7863): 356–364. Bibcode:2021Natur.594..356W. doi:10.1038/s41586-021-03499-y. PMID 34135521. S2CID 235460793.
- 1 2 3 4 Flegontov et al. 2016.
- ↑ Jeong et al. 2019: "the Upper Paleolithic genomes from the Mal’ta and Afontova Gora sites in southern Siberia revealed a genetic profile, often called "Ancient North Eurasians (ANE)", which is deeply related to Paleolithic/Mesolithic hunter-gatherers in Europe"
- ↑ Grebenyuk, Pavel S.; Fedorchenko, Alexander Yu.; Dyakonov, Viktor M.; Lebedintsev, Alexander I.; Malyarchuk, Boris A. (2022). "Ancient Cultures and Migrations in Northeastern Siberia". Humans in the Siberian Landscapes. Springer Geography. Springer International Publishing. p. 93. doi:10.1007/978-3-030-90061-8_4. ISBN 978-3-030-90060-1.
Complete genome sequencing of two individuals from the Yana site has revealed that their genomes belonged to the mitochondrial DNA (mtDNA) haplogroup U, typical among the ancient West Eurasian hunter-gatherers. Analysis of the paternally inherited Y-chromosomes (Y-DNA) has shown that the individuals studied belonged to the haplogroup P1, ancestral to the Q and R haplogroups, widespread among present-day Eurasians and Native Americans (Sikora et al. 2019). The study has revealed that the Ancient North Siberian component of the genome of the Yana individuals ("Ancient North Siberians") was ancestral concerning the "Ancient North Eurasian" component which had been previously found in the genome of the child from the burial at Malta site in the Angara region (Raghavan et al. 2014)
- ↑ Villalba-Mouco, Vanessa; van de Loosdrecht, Marieke S.; Rohrlach, Adam B.; Fewlass, Helen; Talamo, Sahra; Yu, He; Aron, Franziska; Lalueza-Fox, Carles; Cabello, Lidia; Cantalejo Duarte, Pedro; Ramos-Muñoz, José; Posth, Cosimo; Krause, Johannes; Weniger, Gerd-Christian; Haak, Wolfgang (2023-03-01). "A 23,000-year-old southern Iberian individual links human groups that lived in Western Europe before and after the Last Glacial Maximum". Nature Ecology & Evolution. 7 (4): 597–609. Bibcode:2023NatEE...7..597V. doi:10.1038/s41559-023-01987-0. ISSN 2397-334X. PMC 10089921. PMID 36859553.
This is because the ancestry found in Mal'ta and Afontova Gora individuals (Ancient North Eurasian ancestry) received ancestry from UP East Asian/Southeast Asian populations54, who then contributed substantially to EHG55.
- ↑ Sikora, Martin; Pitulko, Vladimir V.; Sousa, Vitor C.; Allentoft, Morten E.; Vinner, Lasse; Rasmussen, Simon; Margaryan, Ashot; de Barros Damgaard, Peter; de la Fuente, Constanza; Renaud, Gabriel; Yang, Melinda A.; Fu, Qiaomei; Dupanloup, Isabelle; Giampoudakis, Konstantinos; Nogués-Bravo, David (June 2019). "The population history of northeastern Siberia since the Pleistocene". Nature. 570 (7760): 182–188. Bibcode:2019Natur.570..182S. doi:10.1038/s41586-019-1279-z. hdl:1887/3198847. ISSN 1476-4687. PMID 31168093. S2CID 174809069.
Using admixture graphs (qpGraph) and outgroup-based estimation of mixture proportions (qpAdm), we find that Yana can be modelled as EWE with ~25% contribution from EEA (Figure 2; Extended Data Fig. 3; Supplementary Information 6).
- ↑ Vallini et al. 2022, Supplementary Information, p. 17: "Paleolithic Siberian populations younger than 40 ky are consistently described as a mix of European and East Asian ancestries,..., however the East Asian component of siberians needs to pick ancestry from different positions along the IUP branch".
- ↑ Lipson, Mark; Reich, David (2017-01-10). "A working model of the deep relationships of diverse modern human genetic lineages outside of Africa". Molecular Biology and Evolution. 34 (4): 889–902. doi:10.1093/molbev/msw293. ISSN 0737-4038. PMC 5400393. PMID 28074030.
- 1 2 Posth, Cosimo; Nakatsuka, Nathan; Lazaridis, Iosif; Skoglund, Pontus; Mallick, Swapan; Lamnidis, Thiseas C.; Rohland, Nadin; Nägele, Kathrin; Adamski, Nicole; Bertolini, Emilie; Broomandkhoshbacht, Nasreen; Cooper, Alan; Culleton, Brendan J.; Ferraz, Tiago; Ferry, Matthew (November 2018). "Reconstructing the Deep Population History of Central and South America". Cell. 175 (5): 1185–1197.e22. doi:10.1016/j.cell.2018.10.027. ISSN 0092-8674. PMC 6327247. PMID 30415837.
- 1 2 Allentoft, Morten E.; Sikora, Martin; Refoyo-Martínez, Alba; Irving-Pease, Evan K.; Fischer, Anders; Barrie, William; Ingason, Andrés; Stenderup, Jesper; Sjögren, Karl-Göran; Pearson, Alice; Sousa da Mota, Bárbara; Schulz Paulsson, Bettina; Halgren, Alma; Macleod, Ruairidh; Jørkov, Marie Louise Schjellerup (January 2024). "Population genomics of post-glacial western Eurasia". Nature. 625 (7994): 301–311. doi:10.1038/s41586-023-06865-0. ISSN 1476-4687. PMID 38200295.
- 1 2 Raff, Jennifer (8 February 2022). Origin: A Genetic History of the Americas. Grand Central Publishing. p. 188. ISBN 978-1-5387-4970-8. "Current estimates suggest that approximately 63% of the First Peoples' ancestry comes from the East Asian group and the rest from the Ancient North Siberians. We're not sure where this interaction took place. Some archaeologists believe that it occurred in East Asia, suggesting that this is where the Siberians moved during the LGM" [...] "There's also a case to be made for this interaction having taken place bear the Lake Baikal region in Siberia from genetic evidence, too" [...] "But other archaeologists and geneticists argue that the meeting of the two grandparent populations of Native Americans occurred because people moved north, not south, in response to the LGM"
- ↑ Reich 2018, p. 81
- 1 2 3 4 Anthony & Brown 2019, pp. 104–106.
- ↑ Willerslev, Eske; Meltzer, David J. (June 2021). "Peopling of the Americas as inferred from ancient genomics". Nature. 594 (7863): 356–364. Bibcode:2021Natur.594..356W. doi:10.1038/s41586-021-03499-y. ISSN 1476-4687. PMID 34135521. S2CID 235460793.
- 1 2 Sikora, Martin; Pitulko, Vladimir V.; Sousa, Vitor C.; Allentoft, Morten E.; Vinner, Lasse; Rasmussen, Simon; Margaryan, Ashot; de Barros Damgaard, Peter; de la Fuente, Constanza; Renaud, Gabriel; Yang, Melinda A.; Fu, Qiaomei; Dupanloup, Isabelle; Giampoudakis, Konstantinos; Nogués-Bravo, David (June 2019). "The population history of northeastern Siberia since the Pleistocene". Nature. 570 (7760): 182–188. Bibcode:2019Natur.570..182S. doi:10.1038/s41586-019-1279-z. hdl:1887/3198847. ISSN 1476-4687. PMID 31168093. S2CID 174809069.
- ↑ "Hall 11 panorama". State Hermitage Museum.
- ↑ Lbova, Liudmila (2021). "The Siberian Paleolithic site of Mal'ta: a unique source for the study of childhood archaeology". Evolutionary Human Sciences. 3: 8, Fig. 6-1. doi:10.1017/ehs.2021.5. PMC 10427291. PMID 37588521. S2CID 231980510.
- ↑ Kozintsev, A. G. (2020-12-31). "The Origin of the Okunev Population, Southern Siberia: The Evidence of Physical Anthropology and Genetics". Archaeology, Ethnology & Anthropology of Eurasia. 48 (4): 135–145. doi:10.17746/1563-0110.2020.48.4.135-145. ISSN 1563-0110.
Recent genetic discoveries call into question the unilinear west vs. east dichotomy (Caucasoid vs. Mongoloid in traditional terms). The actual pattern of differentiation in Eurasia and America proved much more complex.
- ↑ Fu, Qiaomei; Posth, Cosimo; Hajdinjak, Mateja; Petr, Martin; Mallick, Swapan; Fernandes, Daniel; Furtwängler, Anja; Haak, Wolfgang; Meyer, Matthias; Mittnik, Alissa; Nickel, Birgit; Peltzer, Alexander; Rohland, Nadin; Slon, Viviane; Talamo, Sahra (June 2016). "The genetic history of Ice Age Europe". Nature. 534 (7606): 200–205. Bibcode:2016Natur.534..200F. doi:10.1038/nature17993. hdl:10211.3/198594. ISSN 1476-4687. PMC 4943878. PMID 27135931.
- ↑ Maier, Robert; Flegontov, Pavel; Flegontova, Olga; Işıldak, Ulaş; Changmai, Piya; Reich, David (2023-04-14). Nordborg, Magnus; Przeworski, Molly; Balding, David; Wiuf, Carsten (eds.). "On the limits of fitting complex models of population history to f-statistics". eLife. 12: e85492. doi:10.7554/eLife.85492. ISSN 2050-084X. PMC 10310323. PMID 37057893.
- ↑ Zhang X, Ji X, Li C, Yang T, Huang J, Zhao Y, et al. (July 2022). "A Late Pleistocene human genome from Southwest China". Current Biology. 32 (14): 3095–3109.e5. doi:10.1016/j.cub.2022.06.016. PMID 35839766. S2CID 250502011. "In addition to the earliest southern settlement of AMHs in East Asia, ancient migration (40–18 kya) into East Asia via the “Northern Route” from West Eurasia was previously proposed. The “Northern Route” hypothesis would also explain where the subtle shared ancient north Eurasian (ANE) ancestry came from that is then also shared with Native Americans."
- ↑ Svensson, Emma; Günther, Torsten; Hoischen, Alexander; Hervella, Montserrat; Munters, Arielle R.; Ioana, Mihai; Ridiche, Florin; Edlund, Hanna; van Deuren, Rosanne C.; Soficaru, Andrei; de-la-Rua, Concepción; Netea, Mihai G.; Jakobsson, Mattias (2021-07-26). "Genome of Peştera Muierii skull shows high diversity and low mutational load in pre-glacial Europe". Current Biology. 31 (14): 2973–2983.e9. doi:10.1016/j.cub.2021.04.045. hdl:10810/52864. ISSN 0960-9822. PMID 34010592. S2CID 234793812.
- 1 2 Villalba-Mouco, Vanessa; van de Loosdrecht, Marieke S.; Rohrlach, Adam B.; Fewlass, Helen; Talamo, Sahra; Yu, He; Aron, Franziska; Lalueza-Fox, Carles; Cabello, Lidia; Cantalejo Duarte, Pedro; Ramos-Muñoz, José; Posth, Cosimo; Krause, Johannes; Weniger, Gerd-Christian; Haak, Wolfgang (April 2023). "A 23,000-year-old southern Iberian individual links human groups that lived in Western Europe before and after the Last Glacial Maximum". Nature Ecology & Evolution. 7 (4): 597–609. Bibcode:2023NatEE...7..597V. doi:10.1038/s41559-023-01987-0. hdl:10230/57143. ISSN 2397-334X. PMC 10089921. PMID 36859553.
Currently, the strongest affinity to Tianyuan in Holocene European HGs was reported for Eastern European HGs (EHG). This is because the ancestry found in Mal'ta and Afontova Gora individuals (Ancient North Eurasian ancestry) received ancestry from UP East Asian/Southeast Asian populations54, who then contributed substantially to EHG55.
- 1 2 Jeong et al. 2019.
- 1 2 3 Raghavan et al. 2013.
- ↑ Balter 2013.
- ↑ Yang, Melinda A. (2022-01-06). "A genetic history of migration, diversification, and admixture in Asia". Human Population Genetics and Genomics. 2 (1): 1–32. doi:10.47248/hpgg2202010001. ISSN 2770-5005.
- ↑ Sikora, Martin; Pitulko, Vladimir V.; Sousa, Vitor C.; Allentoft, Morten E.; Vinner, Lasse; Rasmussen, Simon; Margaryan, Ashot; de Barros Damgaard, Peter; de la Fuente, Constanza; Renaud, Gabriel; Yang, Melinda A. (June 2019). "The population history of northeastern Siberia since the Pleistocene". Nature. 570 (7760): 182–188. Bibcode:2019Natur.570..182S. doi:10.1038/s41586-019-1279-z. hdl:1887/3198847. ISSN 1476-4687. PMID 31168093. S2CID 174809069.
- 1 2 Lipson, Mark; Reich, David (April 2017). "A Working Model of the Deep Relationships of Diverse Modern Human Genetic Lineages Outside of Africa". Molecular Biology and Evolution. 34 (4): 889–902. doi:10.1093/molbev/msw293. ISSN 0737-4038. PMC 5400393. PMID 28074030.
- ↑ Yang, Melinda A.; Fan, Xuechun; Sun, Bo; Chen, Chungyu; Lang, Jianfeng; Ko, Ying-Chin; Tsang, Cheng-hwa; Chiu, Hunglin; Wang, Tianyi; Bao, Qingchuan; Wu, Xiaohong; Hajdinjak, Mateja; Ko, Albert Min-Shan; Ding, Manyu; Cao, Peng (2020-07-17). "Ancient DNA indicates human population shifts and admixture in northern and southern China". Science. 369 (6501): 282–288. Bibcode:2020Sci...369..282Y. doi:10.1126/science.aba0909. ISSN 0036-8075. PMID 32409524. S2CID 218649510.
- ↑ Yang, Melinda A.; Gao, Xing; Theunert, Christoph; Tong, Haowen; Aximu-Petri, Ayinuer; Nickel, Birgit; Slatkin, Montgomery; Meyer, Matthias; Pääbo, Svante; Kelso, Janet; Fu, Qiaomei (2017-10-23). "40,000-Year-Old Individual from Asia Provides Insight into Early Population Structure in Eurasia". Current Biology. 27 (20): 3202–3208.e9. doi:10.1016/j.cub.2017.09.030. ISSN 0960-9822. PMC 6592271. PMID 29033327.
- ↑ Childebayeva, Ainash; et al. (October 1, 2023). "Bronze Age Northern Eurasian Genetics in the Context of Development of Metallurgy and Siberian Ancestry". BioRxiv: Figure 6. doi:10.1101/2023.10.01.560195. S2CID 263672903.
- 1 2 Grebenyuk, Pavel S.; Fedorchenko, Alexander Yu.; Dyakonov, Viktor M.; Lebedintsev, Alexander I.; Malyarchuk, Boris A. (2022), Bocharnikov, Vladimir N.; Steblyanskaya, Alina N. (eds.), "Ancient Cultures and Migrations in Northeastern Siberia", Humans in the Siberian Landscapes: Ethnocultural Dynamics and Interaction with Nature and Space, Springer Geography, Cham: Springer International Publishing, pp. 89–133, doi:10.1007/978-3-030-90061-8_4, ISBN 978-3-030-90061-8, retrieved 2023-01-09
- ↑ Maier et al. 2023.
- ↑ "Figurine of a woman with the "Spanish rose" hairdo". Art of Mal'ta. Novosibirsk State University.
- ↑ Cassidy, Jim; Ponkratova, Irina; Fitzhugh, Ben, eds. (2022). Maritime Prehistory of Northeast Asia. The Archaeology of Asia-Pacific Navigation. Vol. 6. doi:10.1007/978-981-19-1118-7. ISBN 978-981-19-1117-0. S2CID 249355636.
- 1 2 3 Lazaridis et al. 2016.
- 1 2 Essel, Elena (3 May 2023). "Ancient human DNA recovered from a Palaeolithic pendant". Nature. 618 (7964): 328–332. Bibcode:2023Natur.618..328E. doi:10.1038/s41586-023-06035-2. PMC 10247382. PMID 37138083.
Application of the method to an Upper Palaeolithic deer tooth pendant from Denisova Cave, Russia, resulted in the recovery of ancient human and deer mitochondrial genomes, which allowed us to estimate the age of the pendant at approximately 19,000–25,000 years. Nuclear DNA analysis identifies the presumed maker or wearer of the pendant as a female individual with strong genetic affinities to a group of Ancient North Eurasian individuals who lived around the same time but were previously found only further east in Siberia. Our work redefines how cultural and genetic records can be linked in prehistoric archaeology. (...) Nuclear DNA analysis identifies the presumed maker or wearer of the pendant as a female individual with strong genetic affinities to a group of Ancient North Eurasian individuals who lived around the same time but were previously found only further east in Siberia.
- 1 2 3 4 5 Haak et al. 2015.
- 1 2 Kozintsev, A. G. (2020-12-31). "The Origin of the Okunev Population, Southern Siberia: The Evidence of Physical Anthropology and Genetics". Archaeology, Ethnology & Anthropology of Eurasia. 48 (4): 135–145. doi:10.17746/1563-0110.2020.48.4.135-145. ISSN 1563-0110.
- ↑ Moreno-Mayar et al. 2018.
- 1 2 Wong et al. 2017.
- 1 2 Günther et al. 2018.
- ↑ Gabidullina, L.R.; et al. (2023). "Genetic History of Eurasia Before the Common Era". Opera Medica et Physiologica. 10 (3): 95–117. doi:10.24412/2500-2295-2023-3-95-117.
- 1 2 3 Posth, Cosimo; Yu, He; Ghalichi, Ayshin; Rougier, Hélène; Crevecoeur, Isabelle; Huang, Yilei; Ringbauer, Harald; Rohrlach, Adam B.; Nägele, Kathrin; Villalba-Mouco, Vanessa; Radzeviciute, Rita; Ferraz, Tiago; Stoessel, Alexander; Tukhbatova, Rezeda; Drucker, Dorothée G. (March 2023). "Palaeogenomics of Upper Palaeolithic to Neolithic European hunter-gatherers". Nature. 615 (7950): 117–126. Bibcode:2023Natur.615..117P. doi:10.1038/s41586-023-05726-0. ISSN 1476-4687. PMC 9977688. PMID 36859578.
- ↑ Vallini et al. 2022: "The lack of West Eurasian components in Tianyuan and in subsequent East Asian individuals may provide clues on the resistance of those groups to the incoming UP population movements, or on subsequent reexpansion from a genetically IUP-like population reservoir."
- ↑ Raff 2022, p. 188: "Mal'ta's population -- the ancient Northern Siberians, seems to have encountered the daughter East Asian population described at the beginning of this chapter around 25,000 years ago and interbred with them."..."We're not sure where this interaction took place. Some archaeologists believe that it occurred in East Asia, suggesting that this is where the Siberians moved during the LGM"..."There's also a case to be made for this interaction having taken place bear the Lake Baikal region in Siberia from genetic evidence, too. The ancient Paleo-Siberians, as mentioned earlier in this chapter, split from the East Asian ancestors of Native Americans by about 25,000 years ago. They are known to us from the genomes of an Upper Paleolithic person from the Lake Baikal region known to us from the genomes of an Upper Paleolithic from the Lake Baikal region known as UKY and a person from Northeastern Siberia dating to about 9,800 years ago known as Kolyma1."
- ↑ Raff 2022, pp. 188–189: "But other archaeologists and geneticists argue that the meeting of the two grandparent populations of Native Americans occurred because people moved north, not south, in response to the LGM. In this scenario, Paleo-Siberian descendants, like UKY, could have been the result of a southward repopulation of Siberia out of Beringia. The reason for this is because both mitochondrial and nuclear genomes of Native Americans show that they had been isolated from all other populations for a prolonged period of time, during which they developed the genetic traits found only in Native American populations. This finding, initially based on classical genetic markers and mitochondrial evidence, came to be known as the Beringian Incubation, the Beringian Pause or the Beringian Standstill hypothesis."
- ↑ Grebenyuk, Pavel S.; Fedorchenko, Alexander Yu.; Dyakonov, Viktor M.; Lebedintsev, Alexander I.; Malyarchuk, Boris A. (2022). "Ancient Cultures and Migrations in Northeastern Siberia". Humans in the Siberian Landscapes. Springer Geography. Springer International Publishing. p. 96. doi:10.1007/978-3-030-90061-8_4. ISBN 978-3-030-90060-1.
According to the latest paleogenetic data, East Asian populations migrated to Northeastern Siberia ca. 20,000–18,000 cal BP. The migration was accompanied by their mixing with the descendants of the "Ancient North Siberians", represented by the genome from the Yana and Malta individuals. These processes were reflected in the Beringian tradition's wide proliferation in the region and led to the emergence of several ancestral lineages (Fig. 1) in Extreme Northeastern Asia: the Ancient Paleosiberian population represented by the genome of the individual from Duvanny Yar, and the ancestral Native Americans. The latter type subsequently divided into the Ancient Beringians and all other Native Americans (Moreno-Mayar et al. 2018; Sikora et al. 2019).
- ↑ Hoffecker, John F. (4 March 2016). "Beringia and the global dispersal of modern humans: Beringia and the Global Dispersal of Modern Humans". Evolutionary Anthropology: Issues, News, and Reviews. 25 (2): 64–78. doi:10.1002/evan.21478. PMID 27061035. S2CID 3519553. "The pattern observed in the mtDNA data by Tamm and colleagues has yet to emerge with equal clarity in either Y-DNA or autosomal DNA. The lack of a similar pattern in the Y-DNA data conceivably reflects different histories of the paternal and maternal lineages—a pattern found in other parts of the world (e.g., southern Asia).33 Some support for the standstill hypothesis in autosomal DNA may be found in the distribution of an allele at a microsatellite locus on chromosome 9 (D9S1120) which exhibits a pattern similar to that seen in the mtDNA data.34"
- ↑ Moreno-Mayar et al. 2018.
- ↑ Gakuhari, Takashi; Nakagome, Shigeki; Rasmussen, Simon; Allentoft, Morten E. (25 August 2020). "Ancient Jomon genome sequence analysis sheds light on migration patterns of early East Asian populations". Communications Biology. 3 (1): Fig.1 A, C. doi:10.1038/s42003-020-01162-2. ISSN 2399-3642. PMC 7447786. PMID 32843717.
- ↑ Yu et al. 2020.
- ↑ Osada & Kawai 2021.
- ↑ Cooke, Niall P.; Mattiangeli, Valeria; Cassidy, Lara M.; Okazaki, Kenji; Stokes, Caroline A.; Onbe, Shin; Hatakeyama, Satoshi; Machida, Kenichi; Kasai, Kenji; Tomioka, Naoto; Matsumoto, Akihiko (2021). "Ancient genomics reveals tripartite origins of Japanese populations". Science Advances. 7 (38): eabh2419. Bibcode:2021SciA....7.2419C. doi:10.1126/sciadv.abh2419. PMC 8448447. PMID 34533991. "This affinity is still detectable even if we replace these reference populations with the other Southeast and East Asians (table S6), supporting gene flow between the ancestors of Jomon and Ancient North Siberians, a population widespread in North Eurasia before the LGM (19)."
- ↑ Wang, Ke; Yu, He; Radzevičiūtė, Rita; Kiryushin, Yuriy F.; Tishkin, Alexey A.; Frolov, Yaroslav V.; Stepanova, Nadezhda F.; Kiryushin, Kirill Yu; Kungurov, Artur L.; Shnaider, Svetlana V.; Tur, Svetlana S.; Tiunov, Mikhail P.; Zubova, Alisa V.; Pevzner, Maria; Karimov, Timur (2023-01-12). "Middle Holocene Siberian genomes reveal highly connected gene pools throughout North Asia". Current Biology. 33 (3): 423–433. doi:10.1016/j.cub.2022.11.062. ISSN 0960-9822. PMID 36638796. S2CID 255750546.
- ↑ Kozintsev, A. G. (4 January 2022). "Patterns in the Population History of Northern Eurasia from the Mesolithic to the Early Bronze Age, Based on Craniometry and Genetics". Archaeology, Ethnology & Anthropology of Eurasia. 49 (4): 141. doi:10.17746/1563-0110.2021.49.4.140-151.
Modern Old World groups closest to native Americans in this respect are Chukchi, Koryaks, Kets, and Selkups, and among the ancient groups, Okunev people and those of the Botai culture*, suggesting that both these groups are direct descendants of the Malta-Buret people.
- ↑ Narasimhan 2019.
- ↑ Mattila, Tiina M.; Svensson, Emma M.; Juras, Anna; Günther, Torsten; Kashuba, Natalija; Ala-Hulkko, Terhi; Chyleński, Maciej; McKenna, James; Pospieszny, Łukasz; Constantinescu, Mihai; Rotea, Mihai; Palincaș, Nona; Wilk, Stanisław; Czerniak, Lech; Kruk, Janusz (2023-08-09). "Genetic continuity, isolation, and gene flow in Stone Age Central and Eastern Europe". Communications Biology. 6 (1): 1–13. doi:10.1038/s42003-023-05131-3. hdl:10852/104260. ISSN 2399-3642. PMID 37558731.
- 1 2 3 Zhang, Fan (November 2021). "The genomic origins of the Bronze Age Tarim Basin mummies". Nature. 599 (7884): 256–261. Bibcode:2021Natur.599..256Z. doi:10.1038/s41586-021-04052-7. ISSN 1476-4687. PMC 8580821. PMID 34707286.
The Tarim mummies are among only a few known Holocene populations that derive the majority of their ancestry from Pleistocene ANE groups, who once made up the huntergatherer populations of southern Siberia, and which are represented by individual genomes from the archaeological sites of Mal'ta (MA-1)29 and Afontova Gora (AG3). (...) The Tarim mummies are currently the best representative of the pre-pastoralist ANE-related population that once inhabited Central Asia and southern Siberia (Extended Data Fig. 2A), even though Tarim_EMBA1 postdates these populations in time.
- ↑ Jeong et al. 2020.
- ↑ Zhang, Fan (November 2021). "The genomic origins of the Bronze Age Tarim Basin mummies". Nature. 599 (7884): 256–261. Bibcode:2021Natur.599..256Z. doi:10.1038/s41586-021-04052-7. ISSN 1476-4687. PMC 8580821. PMID 34707286.
To understand this mixed genetic profile, we used qpAdm to explore admixture models of the Dzungarian groups with Tarim_EMBA1 or a terminal Pleistocene individual (AG3) from the Siberian site of Afontova Gora31, as a source (Supplementary Data 1D). AG3 is a distal representative of the ANE ancestry and shows a high affinity with Tarim_EMBA1.
- ↑ Zhang, Fan (November 2021). "The genomic origins of the Bronze Age Tarim Basin mummies". Nature. 599 (7884): 256–261. Bibcode:2021Natur.599..256Z. doi:10.1038/s41586-021-04052-7. ISSN 1476-4687. PMC 8580821. PMID 34707286.
Using qpAdm, we modelled the Tarim Basin individuals as a mixture of two ancient autochthonous Asian genetic groups: the ANE, represented by an Upper Palaeolithic individual from the Afontova Gora site in the upper Yenisei River region of Siberia (AG3) (about 72%), and ancient Northeast Asians, represented by Baikal_EBA (about 28%)
- ↑ Lazaridis, Iosif; Patterson, Nick; Mittnik, Alissa (September 2014). "Ancient human genomes suggest three ancestral populations for present-day Europeans". Nature. 513 (7518): 409–413. arXiv:1312.6639. Bibcode:2014Natur.513..409L. doi:10.1038/nature13673. ISSN 1476-4687. PMC 4170574. PMID 25230663.
Most present-day Europeans derive from at least three highly differentiated populations: west European hunter-gatherers, who contributed ancestry to all Europeans but not to Near Easterners; ancient north Eurasians related to Upper Palaeolithic Siberians, who contributed to both Europeans and Near Easterners; and early European farmers, who were mainly of Near Eastern origin but also harboured west European hunter-gatherer related ancestry.
- ↑ Lazaridis, Patterson & Mittnik 2014.
- ↑ Wang 2019.
- ↑ Mathieson et al. 2018.
- ↑ Lazaridis et al. 2014.
- ↑ Mathieson 2015.
- ↑ Mittnik 2018.
- ↑ Jeong et al. 2018.
- ↑ Raghavan & Skoglund et al. 2014.
- ↑ Fu, Qiaomei; Posth, Cosimo; et al. (May 2, 2016). "The genetic history of Ice Age Europe". Nature. 504 (7606): 200–5. Bibcode:2016Natur.534..200F. doi:10.1038/nature17993. hdl:10211.3/198594. PMC 4943878. PMID 27135931.
- ↑ Raghavan, Maanasa; Skoglund, Pontus; Graf, Kelly E.; Metspalu, Mait; Albrechtsen, Anders; Moltke, Ida; Rasmussen, Simon; Stafford, Thomas W.; Orlando, Ludovic; Metspalu, Ene; Karmin, Monika; Tambets, Kristiina; Rootsi, Siiri; Mägi, Reedik; Campos, Paula F.; Balanovska, Elena; Balanovsky, Oleg; Khusnutdinova, Elza; Litvinov, Sergey; Osipova, Ludmila P.; Fedorova, Sardana A.; Voevoda, Mikhail I.; DeGiorgio, Michael; Sicheritz-Ponten, Thomas; Brunak, Søren; Demeshchenko, Svetlana; Kivisild, Toomas; Villems, Richard; Nielsen, Rasmus; Jakobsson, Mattias; Willerslev, Eske (2 January 2014). "Upper Palaeolithic Siberian genome reveals dual ancestry of NativeAmericans". Nature. 505 (7481): 87–91. Bibcode:2014Natur.505...87R. doi:10.1038/nature12736. ISSN 0028-0836. PMC 4105016. PMID 24256729. "MA-1 individual was predicted as having relatively dark pigmentation for hair, skin and eyes. However, we note that this analysis is based on only 17 SNPs at low depth-of-coverage, and even with the full set of SNPs the method has limited prediction accuracy. [...] MA-1 has the G allele at rs6152 that is associated with high risk for male pattern baldness. As expected, MA-1 also carries the ancestral C allele at rs4988235 [...] It does not have the allele associated with hair thickness and shovel-shaped incisors in Asians (rs3827760), while the position that is associated with dry earwax in East Asians (rs17822931) is not covered in the draft genome data." }
- 1 2 3 Mathieson et al. 2018 "Supplementary Information page 52: "The derived allele of the KITLG SNP rs12821256 that is associated with – and likely causal for blond hair in Europeans is present in one hunter-gatherer from each of Samara, Motala and Ukraine (I0124, I0014 and I1763), as well as several later individuals with Steppe ancestry. Since the allele is found in populations with EHG but not WHG ancestry, it suggests that its origin is in the Ancient North Eurasian (ANE) population. Consistent with this, we observe that the earliest known individual with the derived allele (supported by two reads) is the ANE individual Afontova Gora 3, which is directly dated to 16130-15749 cal BCE (14710±60 BP, MAMS-27186: a previously unpublished date that we newly report here). We cannot determine the status of rs12821256 in Afontova Gora 2 and MA-1 due to lack of sequence coverage at this SNP."
- ↑ Zhang, Fan; Ning, Chao; Scott, Ashley; Fu, Qiaomei; Bjørn, Rasmus; Li, Wenying; Wei, Dong; Wang, Wenjun; Fan, Linyuan; Abuduresule, Idilisi; Hu, Xingjun (November 2021). "The genomic origins of the Bronze Age Tarim Basin mummies". Nature. 599 (7884): 256–261. Bibcode:2021Natur.599..256Z. doi:10.1038/s41586-021-04052-7. ISSN 1476-4687. PMC 8580821. PMID 34707286.
The Tarim mummies' so-called Western physical features are probably due to their connection to the Pleistocene ANE gene pool
- ↑ "A Craniometric Investigation of The Bronze Age Settlement of Xinjiang - Important | PDF". Scribd. Retrieved 2023-04-04.
The results fail to demonstrate close phenetic affinities between the early inhabitants of Qa¨wrighul and any of the proposed sources for immigrants to the Tarim Basin. The absence of close affinities to outside populations renders it unlikely that the human remains recovered from Qa¨wrighul represent the unadmixed remains of colonists from the Afanasievo or Andronovo cultures of the steppe lands, or inhabitants of the urban centers of the Oxus civilization of Bactria.
- ↑ Carlberg & Hanel 2020, "Interestingly, ancient North Eurasian derived populations, such as eastern hunter-gatherers and Yamnayas, carried the blond hair allele rs12821256 of the KITLG gene to Europe. Its first evidence was described in an 18 000 years old ancient North Eurasian west of Lake Baikal".
- ↑ Hartz, Patricia. "KIT LIGAND; KITLG". Online Mendelian Inheritance in Man.
- ↑ Khan, Razib. "KITLG makes you white skinned?". Discover Magazine.
- ↑ "P21583". Uniprot.
- ↑ Guenther et al. 2014.
- ↑ Evans, Gavin (2019). Skin Deep: Dispelling the Science of Race (1 ed.). Simon and Schuster. p. 139. ISBN 9781786076236.| "Japanese research in 2006 found that the genetic mutation that prompted the evolution of blond hair dates to the ice age that happened around 11,000 years ago. Since then, the 17,000-year-old remains of a blond- haired North Eurasian hunter-gatherer have been found in eastern Siberia, suggesting an earlier origin."
- ↑ Reich, David (2018). Who We are and How We Got Here: Ancient DNA and the New Science of the Human Past. Oxford University Press. ISBN 978-0198821250. "The earliest known example of the classic European blond hair mutation is in an Ancient North Eurasian from the Lake Baikal region of eastern Siberia from seventeen thousand years ago. The hundreds of millions of copies of this mutation in central and western Europe today likely derive from a massive migration of people bearing Ancient North Eurasian ancestry, an event that is related in the next chapter."
- ↑ Carlberg & Hanel 2020, "Interestingly, ancient North Eurasian derived populations, such as eastern hunter-gatherers and Yamnayas, carried the blond hair allele rs12821256 of the KITLG gene to Europe. Its first evidence was described in an 18 000 years old ancient North Eurasian west of Lake Baikal.".
- ↑ Zhang, Fan (November 2021). "The genomic origins of the Bronze Age Tarim Basin mummies". Nature. 599 (7884): 256–261. Bibcode:2021Natur.599..256Z. doi:10.1038/s41586-021-04052-7. ISSN 1476-4687. PMC 8580821. PMID 34707286.
- ↑ "The Mal'ta - Buret' venuses and culture in Siberia". donsmaps.com.
Notes
- ↑ Vallini et al. (2022) created a TreeMix model in which "admixture events in varying proportions between sister groups of Kostenki14 and Tianyuan is supported, and can indeed explain this observation (fig. 1A, purple leaves; Supplementary fig. S5 and Section 3.6, Supplementary Material online)".
- ↑ Sikora et al. (2019) model the Yana individuals as 22% East Eurasian and the remainder West Eurasian ("Using admixture graphs and outgroup-based estimation of mixture proportions, we find that ANS can be modelled as early West Eurasian with an approximately 22% contribution from early East Asians"). Massilani et al. (2020) model the Yana individuals as around one-third East Eurasian and two-thirds West Eurasian.Vallini et al. (2022) model Yana as 50% West Eurasian and 50% East Eurasian.
- ↑ Vallini et al. (2022) created a TreeMix model in which "admixture events in varying proportions between sister groups of Kostenki14 and Tianyuan is supported, and can indeed explain this observation (fig. 1A, purple leaves; Supplementary fig. S5 and Section 3.6, Supplementary Material online)".
- ↑ Sikora et al. (2019) model the Yana individuals as 22% East Eurasian and the remainder West Eurasian. Massilani et al. (2020) model the Yana individuals as around one-third East Eurasian and two-thirds West Eurasian.Vallini et al. (2022) model Yana as 50% West Eurasian and 50% East Eurasian.
- ↑ "Recent paleogenomic studies have shown that migrations of Western steppe herders (WSH) beginning in the Eneolithic (ca. 3300–2700 BCE) profoundly transformed the genes and cultures of Europe and central Asia... The migration of these Western steppe herders (WSH), with the Yamnaya horizon (ca. 3300–2700 BCE) as their earliest representative, contributed not only to the European Corded Ware culture (ca. 2500–2200 BCE) but also to steppe cultures located between the Caspian Sea and the Altai-Sayan mountain region, such as the Afanasievo (ca. 3300–2500 BCE) and later Sintashta (2100–1800 BCE) and Andronovo (1800–1300 BCE) cultures."[75]
Bibliography
- Anthony, David W.; Brown, Dorcas R. (2019). "Late Bronze Age midwinter dog sacrifices and warrior initiations at Krasnosamarskoe, Russia". In Olsen, Birgit A.; Olander, Thomas; Kristiansen, Kristian (eds.). Tracing the Indo-Europeans: New evidence from archaeology and historical linguistics. Oxbow Books. ISBN 978-1-78925-273-6.
- Balter, M. (25 October 2013). "Ancient DNA Links Native Americans With Europe". Science. 342 (6157): 409–410. Bibcode:2013Sci...342..409B. doi:10.1126/science.342.6157.409. PMID 24159019.
- Dolitsky, Alexander B.; Ackerman, Robert E.; Aigner, Jean S.; et al. (June 1985). "Siberian Paleolithic Archaeology: Approaches and Analytic Methods [and Comments and Replies]". Current Anthropology. 26 (3): 361–378. doi:10.1086/203280. S2CID 147371671.
- Carlberg, Carsten; Hanel, Andrea (2020). "Skin colour and vitamin D: An update". Experimental Dermatology. 29 (9): 864–875. doi:10.1111/exd.14142. PMID 32621306.
- Flegontov, Pavel; Changmai, Piya; Zidkova, Anastassiya; Logacheva, Maria D.; et al. (11 February 2016). "Genomic study of the Ket: a Paleo-Eskimo-related ethnic group with significant ancient North Eurasian ancestry". Scientific Reports. 6 (1): 20768. arXiv:1508.03097. Bibcode:2016NatSR...620768F. doi:10.1038/srep20768. PMC 4750364. PMID 26865217.
- Günther, Torsten; Malmström, Helena; Svensson, Emma M.; Omrak, Ayça; et al. (9 January 2018). "Population genomics of Mesolithic Scandinavia: Investigating early postglacial migration routes and high-latitude adaptation". PLOS Biology. 16 (1): e2003703. doi:10.1371/journal.pbio.2003703. PMC 5760011. PMID 29315301.
- Haak, Wolfgang; Lazaridis, Iosif; Patterson, Nick; et al. (2 March 2015). "Massive migration from the steppe was a source for Indo-European languages in Europe". Nature. 522 (7555): 207–211. arXiv:1502.02783. Bibcode:2015Natur.522..207H. doi:10.1038/nature14317. PMC 5048219. PMID 25731166.
- Guenther, Catherine A; Tasic, Bosiljka; Luo, Liqun; Bedell, Mary A; Kingsley, David M (July 2014). "A molecular basis for classic blond hair color in Europeans". Nature Genetics. 46 (7): 748–752. doi:10.1038/ng.2991. PMC 4704868. PMID 24880339.
- Jeong, Choongwon; Wilkin, Shevan; Amgalantugs, Tsend; Bouwman, Abigail S.; et al. (27 November 2018). "Bronze Age population dynamics and the rise of dairy pastoralism on the eastern Eurasian steppe". Proceedings of the National Academy of Sciences of the United States of America. 115 (48): E11248–E11255. Bibcode:2018PNAS..11511248J. doi:10.1073/pnas.1813608115. ISSN 0027-8424. PMC 6275519. PMID 30397125.
- Jeong, Choongwon; Balanovsky, Oleg; Lukianova, Elena; Kahbatkyzy, Nurzhibek; et al. (June 2019). "The genetic history of admixture across inner Eurasia". Nature Ecology & Evolution. 3 (6): 966–976. Bibcode:2019NatEE...3..966J. doi:10.1038/s41559-019-0878-2. PMC 6542712. PMID 31036896.
- Jeong, Choongwon; Wang, Ke; Wilkin, Shevan; Taylor, William Timothy Treal; et al. (26 March 2020). "A dynamic 6,000-year genetic history of Eurasia's Eastern Steppe". Cell. 183 (4): 890–904.e29. bioRxiv 10.1101/2020.03.25.008078. doi:10.1016/j.cell.2020.10.015. hdl:21.11116/0000-0007-77BF-D. PMC 7664836. PMID 33157037. S2CID 214725595.
- Kanzawa-Kiriyama, Hideaki; Kryukov, Kirill; Jinam, Timothy A; Hosomichi, Kazuyoshi; et al. (February 2017). "A partial nuclear genome of the Jomons who lived 3000 years ago in Fukushima, Japan". Journal of Human Genetics. 62 (2): 213–221. doi:10.1038/jhg.2016.110. PMC 5285490. PMID 27581845.
- Kurbel, Sven (2019). "Was the Last Ice Age dusty climate instrumental in spreading of the three "Celtic" diseases (hemochromatosis, cystic fibrosis and palmar fibromatosis)?". Medical Hypotheses. 122: 134–138. doi:10.1016/j.mehy.2018.11.006. ISSN 0306-9877. PMID 30593397. S2CID 58661828.
- Lazaridis, Iosif; Nadel, Dani; Rollefson, Gary; et al. (25 July 2016). "Genomic insights into the origin of farming in the ancient Near East". Nature. 536 (7617): 419–424. Bibcode:2016Natur.536..419L. doi:10.1038/nature19310. PMC 5003663. PMID 27459054.
- Lazaridis, Iosif; Patterson, Nick; Mittnik, Alissa; et al. (17 September 2014). "Ancient human genomes suggest three ancestral populations for present-day Europeans". Nature. 513 (7518): 409–413. arXiv:1312.6639. Bibcode:2014Natur.513..409L. doi:10.1038/nature13673. hdl:11336/30563. PMC 4170574. PMID 25230663.
- Maier, Robert; Flegontov, Pavel; Flegontova, Olga; Isildak, Ulas; Changmai, Piya; Reich, David (2023). "On the limits of fitting complex models of population history to f-statistics". eLife. 12: e85492. doi:10.7554/eLife.85492. ISSN 2050-084X. PMC 10310323. PMID 37057893. S2CID 258134572.
- Diyendo Massilani; Laurits Skov; Mateja Hajdinjak; Byambaa Gunchinsuren; Damdinsuren Tseveendorj; Seonbok Yi; Jungeun Lee; Sarah Nagel; Birgit Nickel; Thibaut Devièse; Tom Higham; Matthias Meyer; Janet Kelso; Benjamin M. Peter; Svante Pääbo (2020). "Denisovan ancestry and population history of early East Asians". Science. 370 (6516): 579–583. doi:10.1126/science.abc1166. PMID 33122380. S2CID 225957149.
- Mathieson, Iain; Alpaslan-Roodenberg, Songül; Posth, Cosimo; et al. (21 February 2018). "The genomic history of southeastern Europe" (PDF). Nature. 555 (7695): 197–203. Bibcode:2018Natur.555..197M. doi:10.1038/nature25778. PMC 6091220. PMID 29466330.
- Mathieson, Iain (November 23, 2015). "Genome-wide patterns of selection in 230 ancient Eurasians". Nature. Nature Research. 528 (7583): 499–503. Bibcode:2015Natur.528..499M. doi:10.1038/nature16152. PMC 4918750. PMID 26595274.
- Mittnik, Alisa (January 30, 2018). "The genetic prehistory of the Baltic Sea region". Nature Communications. Nature Research. 9 (1): 442. Bibcode:2018NatCo...9..442M. doi:10.1038/s41467-018-02825-9. PMC 5789860. PMID 29382937.
- Moreno-Mayar, J. Víctor; Vinner, Lasse; de Barros Damgaard, Peter; de la Fuente, Constanza; et al. (7 December 2018). "Early human dispersals within the Americas" (PDF). Science. 362 (6419): eaav2621. Bibcode:2018Sci...362.2621M. doi:10.1126/science.aav2621. PMID 30409807. S2CID 53241760.
- Moreno-Mayar, J. Víctor; Potter, Ben A.; Vinner, Lasse; Steinrücken, Matthias; et al. (January 2018). "Terminal Pleistocene Alaskan genome reveals first founding population of Native Americans" (PDF). Nature. 553 (7687): 203–207. Bibcode:2018Natur.553..203M. doi:10.1038/nature25173. PMID 29323294. S2CID 4454580.
- Narasimhan, Vagheesh M. (September 6, 2019). "The formation of human populations in South and Central Asia". Science. American Association for the Advancement of Science. 365 (6457): eaat7487. bioRxiv 10.1101/292581. doi:10.1126/science.aat7487. PMC 6822619. PMID 31488661.
- Raghavan, Maanasa; Skoglund, Pontus; Graf, Kelly E.; Metspalu, Mait; et al. (20 November 2013). "Upper Palaeolithic Siberian genome reveals dual ancestry of Native Americans". Nature. 505 (7481): 87–91. Bibcode:2014Natur.505...87R. doi:10.1038/nature12736. PMC 4105016. PMID 24256729.
- Raghavan, Maanasa; Skoglund, Pontus; et al. (2014). "Upper Palaeolithic Siberian Genome Reveals Dual Ancestry of Native Americans". Nature. 505 (7481): 87–91. Bibcode:2014Natur.505...87R. doi:10.1038/nature12736. PMC 4105016. PMID 24256729.
- Osada, Naoki; Kawai, Yosuke (2021). "Exploring models of human migration to the Japanese archipelago using genome-wide genetic data". Anthropological Science. 129 (1): 45–58. doi:10.1537/ase.201215.
- Vallini, Leonardo; Marciani, Giulia; Aneli, Serena; Bortolini, Eugenio; et al. (2022). "Genetics and Material Culture Support Repeated Expansions into Paleolithic Eurasia from a Population Hub Out of Africa". Genome Biology and Evolution. 14 (4). doi:10.1093/gbe/evac045. PMC 9021735. PMID 35445261.
- Wang, Chuan-Chao (February 4, 2019). "Ancient human genome-wide data from a 3000-year interval in the Caucasus corresponds with eco-geographic regions Eurasia". Nature Communications. Nature Research. 10 (1): 590. Bibcode:2019NatCo..10..590W. doi:10.1038/s41467-018-08220-8. PMC 6360191. PMID 30713341.
- Wong, Emily H.M.; Khrunin, Andrey; Nichols, Larissa; Pushkarev, Dmitry; et al. (January 2017). "Reconstructing genetic history of Siberian and Northeastern European populations". Genome Research. 27 (1): 1–14. doi:10.1101/gr.202945.115. PMC 5204334. PMID 27965293.
- Yu, He; Spyrou, Maria A.; Karapetian, Marina; Shnaider, Svetlana; et al. (June 2020). "Paleolithic to Bronze Age Siberians Reveal Connections with First Americans and across Eurasia". Cell. 181 (6): 1232–1245.e20. doi:10.1016/j.cell.2020.04.037. PMID 32437661. S2CID 218710761.
| Sub-topics | |
|---|---|
| Genetic history by region | |
| Population genetics by group |
|
| |