| Pterosauria | |
| Kaup, 1834 | |
| Okres istnienia: trias późny–kreda późna | |
 | |
| Systematyka | |
| Domena | |
|---|---|
| Królestwo | |
| Typ | |
| Podtyp | |
| Gromada | |
| Podgromada | |
| Infragromada | |
| (bez rangi) | archozaury? |
| (bez rangi) | Avemetatarsalia? |
| (bez rangi) | pterozauromorfy |
| Rząd |
pterozaury |
Pterozaury, gady latające (Pterosauria, z gr. pteron – skrzydło + sauros – jaszczur) – rząd latających archozauromorfów, prawdopodobnie archozaurów (gadów naczelnych), żyjących od późnego triasu do końca kredy (230 do 65,5 mln lat temu). Najczęściej klasyfikowane jako archozaury z kladu Avemetatarsalia (obejmującego ptaki i archozaury bliżej spokrewnione z nimi niż z krokodylami) blisko spokrewnione z dinozauromorfami[1][2][3]; z niektórych analiz kladystycznych wynika jednak, że mogły być przedstawicielami kladu Archosauriformes nienależącymi do archozaurów[4] lub nawet archozauromorfami nienależącymi do Archosauriformes[5][6].
Były pierwszymi aktywnie latającymi kręgowcami i największymi latającymi zwierzętami odkrytymi przez człowieka. Przez 150 milionów lat, zanim wymarły, dominowały w powietrzu, skolonizowały wszystkie kontynenty i wykształciły wielką rozmaitość form i rozmiarów. Do dzisiaj odkryto ok. 60 rodzajów i 120 gatunków pterozaurów począwszy od wielkości wróbla do rozpiętości skrzydeł powyżej 12 metrów (więcej niż myśliwiec F-16).
Historia odkryć
Pterozaury zostały odkryte w 1784 w Solnhofen przez włoskiego uczonego Cosimo Colliniego, który sądził, że były zwierzętami pływającymi, a nie latającymi[7]. W XIX wieku Georges Cuvier uznał, że latały[8].
Pierwsze skamieniałości pterozaurów odkryto w późnojurajskich wapieniach niemieckiego Solnhofen w 1784[9]. W 1831 roku Georg August Goldfuss opisał pterozaura Scaphognathus crassirostris z włóknistymi strukturami. Badacz Edward Newman z XIX wieku sądził, że pterozaury były latającymi torbaczami[10]. Większość paleontologów obecnie sądzi, że były przystosowane do aktywnego lotu, a nie tylko szybowały jak uważano dawniej.
Większość skamieniałych szczątków nie zachowała się dobrze. Wiele kości było zniekształconych i potrzeba było żmudnej pracy paleontologów, aby właściwie je poskładać. Najlepiej zachowane kości są znajdywane od 1974 r. w Araripe Plateau, w Brazylii.

Budowa
Pierwsze pterozaury miały w pełni uzębione szczęki i długie ogony, natomiast u form późniejszych nastąpiła redukcja ogona, a szczęki pozbawione zębów upodobniły się bardziej do dzioba. Prawdopodobnie rozwinęły aktywny lot od samego początku, ponieważ nie znaleziono żadnych form przystosowanych jedynie do lotu szybowego polegającego na rzucaniu się z drzew lub skał. Budowa ciała i nogi przypominają małego dwunożnego dinozaura. Chodząc po ziemi, utrzymywały pozycję pionową. Mniejsze prawie na pewno mogły biegać i skakać do lotu, trzepocząc skrzydłami. Największe musiały wznosić się podobnie jak olbrzymie współczesne ptaki: kondory. Ich skrzydła zbudowane były z błony skórnej rozpiętej między wydłużonym czwartym palcem (serdeczny) przedniej kończyny a bokiem ciała podobnie jak dzisiejsze nietoperze. Skrzydła długie i wąskie mogły częściowo składać się w locie, dzięki temu uzyskiwały większą zwrotność, szybkość i sterowność. Ich dobrze rozwinięty mostek utrzymywał silne mięśnie poruszające skrzydłami. Kości podobnie jak u ptaków puste w środku (pneumatyczne) były bardzo lekkie. Miały wielkie ażurowe czaszki, często wydłużone, prawdopodobnie przystosowane do łapania ryb, owadów i innej małej zdobyczy. Kostne grzebienie u niektórych gatunków były prawdopodobnie oznaką dymorfizmu płciowego. Odnalezione kości czaszek wskazują, że miały stosunkowo duże mózgi i duże oczy. Najbardziej rozrośnięte były części mózgu odpowiedzialne za orientację w przestrzeni i koordynację ruchów w czasie lotu. Rogowe dzioby u pierwszych form były wyposażone w ostre zęby, które u niektórych późniejszych form zanikły. Z czasem dzięki zrośnięciu się kręgów w odcinku piersiowym i krzyżowym kręgosłup nabrał sztywności, a tułów uległ skróceniu. Wiele, jeżeli nie wszystkie, miały złączone błoną stopy, które mogły być dowodem umiejętności pływania lub mogły mieć aerodynamiczną funkcję jak we współczesnych latawcach. Obecnie przeważa pogląd, że przynajmniej część musiała być stałocieplna, ponieważ wskazuje na to ich aktywny, wysokoenergetyczny tryb życia i odkryte odciśnięte włoski na niektórych skamieniałościach.
Tryb życia
Poruszanie się
Naukowcy od dawna zadają sobie pytanie, w jakim stopniu były sprawnymi lotnikami. Dawniej sądzono, że były jedynie zdolne do lotu ślizgowego, startując z wysoko położonych miejsc i wykorzystywały wznoszące prądy powietrzne nad zbiornikami wodnymi, jak to robią współczesne albatrosy. Taki sposób latania nie potrzebuje dużych nakładów energii, co jest istotnym argumentem, jeśli zakładać, że były zwierzętami zmiennocieplnymi.

Jednak w ten sposób mogły latać jedynie dość duże gatunki, z dużą powierzchnią skrzydeł. Małe pterozaury musiały szybko poruszać skrzydłami podobnie jak nietoperze, a więc być w stanie wystartować z miejsca i wznosić się bez pomocy wiatru. Z uwagi na to, że do latania potrzeba wysokiego nakładu energii, uważa się dzisiaj, że były zwierzętami stałocieplnymi, czego dodatkowym dowodem mogą być odciśnięte w skałach włosy pokrywające niektóre z nich.
Pojawiła się również teoria, zgodnie z którą wzbijały się do lotu, podskakując na czterech kończynach tak, jak czynią to żaby. Wskazuje na to kilkukrotnie silniejsza budowa skrzydeł od tylnych łap[11].
Także niewyjaśnioną kwestią pozostaje sposób poruszania się pterozaurów na ziemi. Ścierają się tutaj dwie alternatywne teorie. Jedni naukowcy sądzą, że mogły chodzić po ziemi, posługując się nogami w sposób, jak robią to dzisiejsze ptaki. Argumentują tę tezę podobną budową anatomiczną nóg obu grup. Inni zaś skłaniają się ku twierdzeniu, że pterozaury chodziły po ziemi na czterech nogach, a jako drugiej pary nóg używały złożonych wpół do tyłu skrzydlatych ramion, podobnie jak współczesne nietoperze wampirowate. Trzecia teoria zakłada, że pomagały sobie przy chodzeniu przednimi kończynami tylko największe pterozaury.
Pożywienie
Sądząc na podstawie uzębienia, większość była zwierzętami mięsożernymi. Wcześniejsze długoogonowe żyły głównie blisko wybrzeży morskich lub nad innymi większymi wodami i prawdopodobnie odżywiały się głównie rybami i kałamarnicami. Ich dzioby były bardzo wydłużone, wypełnione ostrymi, cienkimi zębami, którymi mogły złapać znad powierzchni wody i przytrzymać śliską rybę. Późniejsze małe pterozaury prawdopodobnie polowały na owady. Można domniemywać, że większe gatunki nie były aktywnymi myśliwymi, a podobnie jak dzisiejsze kondory żywiły się padliną. Były też formy mocno wyspecjalizowane jak niezwykły Pterodaustro z Ameryki Południowej, który miał bardzo długi dziób i cienkie zęby, którymi mógł filtrować wodę morską lub bogate w glony i skorupiaki słone jeziora jak flamingi.
Środowisko życia
Pterozaury opanowały nie tylko wybrzeża, ale na pewno żyły w innych środowiskach lądowych jak lasy, góry lub nawet pustynie. Prawdopodobnie jak współczesne ptaki morskie zbierające się w duże stada na czas lęgów tak i pterozaury mogły tworzyć większe kolonie. Pogląd taki nie jest tylko teoretycznym przypuszczeniem, ale opiera na odkrytym stanowisku na pustyni Atacama w Chile, gdzie na bardzo niewielkiej przestrzeni znaleziono szczątki wielu małych pterozaurów.
Rozmnażanie
Pterozaury były jak wszystkie archozaury zwierzętami jajorodnymi. W 2004 r. opublikowano opis skamieniałego zarodka z dolnej kredy sprzed około 121 milionów lat znalezionego w północno-wschodniej chińskiej prowincji Liaoning. Dobrze zachowane jajo trochę mniejsze od kurzego zachowało się w całości, ponieważ zarodek nie zdążył się wykluć. Prawdopodobnie nastąpiła erupcja wulkanu i dzięki temu jajo tak dobrze się zachowało. Bardzo dobrze zachowany szkielet, a widoczne kości ramion, klatki piersiowej oraz charakterystyczny wydłużony czwarty palec jednoznacznie wskazują na potomka pterozaura. Embrion miał bardzo dobrze rozwinięte skrzydła, co może świadczyć, że mały pterozaur wkrótce po wykluciu był zdolny do samodzielnego lotu. Mógł też być zdolny do samodzielnego zdobywania pożywienia i nie być zależnym od rodziców[12].
Bennett (2018) opisał szkielet młodego osobnika należącego do rodzaju Pteranodon odkrytego w osadach górnokredowej formacji Niobrara (Kansas, Stany Zjednoczone); odkryty szkielet różni się od szkieletów większych osobników wyłącznie rozmiarami. W ocenie autora podobieństwo budowy szkieletu młodych i dorosłych pterozaurów dowodzi, że pterozaury były zagniazdownikami. Szkielety młodych osobników z rodzaju Pteranodon rzadko współwystępują ze szkieletami osobników dorosłych, co może dowodzić, że u tych pterozaurów osobniki młode i dorosłe zasiedlały inne środowiska i zajmowały inne nisze ekologiczne[13].
Ewolucja
Pterozaury były pierwszymi kręgowcami w historii życia na ziemi, które opanowały sztukę aktywnego latania[14]. Oprócz nich udało się to jeszcze ptakom i nietoperzom i choć każda z tych grup rozwijała się niezależnie, to można wskazać na konwergencję podobieństwa między nimi. Pterozaury z wyglądu są jakby skrzyżowaniem późniejszych ptaków i nietoperzy. Tak jak ptaki z czasem zredukowały ogon, wykształciły różnego rodzaju dzioby i niektóre utraciły zęby, a tak jak u nietoperzy elementem nośnym skrzydeł była błona skórna.

W toku ewolucji pterozaury wykształciły dwie linie. Wcześniejsze długoogonowe (Rhamphorhynchoidea) charakteryzowały się długim ogonem i stosunkowo krótkimi kośćmi kończyn przednich i późniejsze krótkoogonowe (Pterodactyloidea) miała z kolei tylko bardzo krótkie ogony i długie kości przednich łap. Obecnie odchodzi się już od tego klasycznego podziału i nie zakłada się, że krótkoogonowe pochodzą od długoogonowych, ale zgodnie z ich filogenezą stanowią grupę parafiletyczną.
Systematyka
Pterozaury tradycyjnie były dzielone na 2 podrzędy: wcześniejszy (od późnego triasu do późnej jury) Rhamphorhynchoidea o długich ogonach jak Dimorphodon i Rhamphorhynchus i późniejsze (od późnej jury do późnej kredy), o krótkich ogonach pterodaktyle (Pterodactyloidea) takie jak Pterodactylus, Pteranodon i ogromny Quetzalcoatlus.
- RZĄD PTEROZAURY (wymarły)
- Podrząd Rhamphorhynchoidea
- Rodzina dimorfodony (Dimorphodontidae)
- Rodzina anurognaty (Anurognathidae)
- Rodzina kampylognatoidy (Campylognathoididae)
- Rodzina ramforynchy (Rhamphorhynchidae)
- Podrząd pterodaktyle (Pterodactyloidea)
- Nadrodzina Ornithocheiroidea
- Rodzina Istiodactylidae
- Rodzina Ornithocheiridae
- Rodzina Pteranodontidae
- Rodzina Nyctosauridae
- Nadrodzina Ctenochasmatoidea
- Rodzina Gallodactylidae
- Rodzina Pterodactylidae
- Rodzina Ctenochasmatidae
- Nadrodzina Dsungaripteroidea
- Rodzina Germanodactylidae
- Rodzina Dsungaripteridae
- Nadrodzina Azhdarchoidea
- Rodzina Lonchodectidae
- Rodzina Tapejaridae
- Rodzina Azhdarchidae
- Nadrodzina Ornithocheiroidea
- Podrząd Rhamphorhynchoidea
Wybrane rodzaje
- Dsungaripterus, rozpiętość skrzydeł 3 metry, niezwykły kostny grzebień wzdłuż dzioba, i długa wąska, zakrzywiona szczęka z wielkimi tępymi zębami do zgniatania skorupiaków. Przód bezzębny zakrzywiony do góry. Żył w dolnej kredzie.
- Pteranodon, miał 1,8 metra długości, rozpiętość skrzydeł 7,5 m, żył w górnej kredzie.
- Pterodactylus, rozpiętość skrzydeł 50 do 75 cm, żył w górnej jurze nad brzegami jezior.
- Pterodaustro, kredowy pterozaur z Ameryki Płd., rozpiętość skrzydeł 2 metry, dziób bardzo długi i łagodnie zakrzywiony w górę wypełniony cienkimi jak szczeciny zębami, których mógł używać do filtrowania pokarmu w płytkiej wodzie jak współczesne flamingi.
- Quetzalcoatlus, rozpiętość skrzydeł 12 metrów, ale ważył tylko 50 kg, żył w górnej kredzie.
- Rhamphorhynchus jurajski pterozaur z rombem na końcu ogona, który mógł mu służyć jako ster podczas lotu.
- Anhanguera piscator
 Dimorphodon
Dimorphodon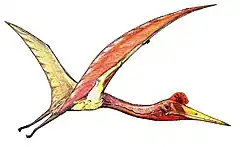 Quetzalcoatlus
Quetzalcoatlus
Przypisy
- ↑ David W.E. Hone, Michael J. Benton. An evaluation of the phylogenetic relationships of the pterosaurs among archosauromorph reptiles. „Journal of Systematic Paleontology”. 5 (4), s. 465–469, 2007. DOI: 10.1017/S1477201907002064. (ang.).
- ↑ Stephen L. Brusatte, Michael J. Benton, Julia B. Desojo, Max C. Langer. The higher-level phylogeny of Archosauria (Tetrapoda: Diapsida). „Journal of Systematic Palaeontology”. 8 (1), s. 3–47, 2010. DOI: 10.1080/14772010903537732. (ang.).
- ↑ Sterling J. Nesbitt. The early evolution of archosaurs. Relationships and the origin of major clades. „Bulletin of the American Museum of Natural History”. 352, s. 1–292, 2011. (ang.).
- ↑ S. Christopher Bennett. The phylogenetic position of the Pterosauria within the Archosauromorpha re-examined. „Historical Biology: An International Journal of Paleobiology”. 25 (5–6), s. 545–563, 2013. DOI: 10.1080/08912963.2012.725727. (ang.).
- ↑ David Peters. A reexamination of four prolacertiforms with implications for pterosaur phylogenesis. „Rivista Italiana di Paleontologia e Stratigrafia”. 106, s. 293–336, 2000. (ang.).
- ↑ Silvio Renesto, Giorgio Binelli. Vallesaurus cenensis Wild, 1991, a drepanosaurid (Reptilia, Diapsida) from the Late Triassic of Northern Italy. „Rivista Italiana di Paleontologia e Stratigrafia”. 112, s. 77–94, 2006. (ang.).
- ↑ Philippe Taquet, Kevin Padian, The earliest known restoration of a pterosaur and the philosophical origins of Cuvier’s Ossemens Fossiles, „Comptes Rendus Palevol”, 3 (2), 2004, s. 157–175, DOI: 10.1016/j.crpv.2004.02.002, ISSN 1631-0683 [dostęp 2024-01-06].
- ↑ Mohamed Y. Zakaria, Haithem E. Taha, Muhammad R. Hajj, Design Optimization of Flapping Ornithopters: The Pterosaur Replica in Forward Flight, „Journal of Aircraft”, 53 (1), 2016, s. 48–59, DOI: 10.2514/1.C033154, ISSN 0021-8669 [dostęp 2024-01-07] (ang.).
- ↑ David M. Martill i inni, A new pterodactyloid pterosaur with a unique filter-feeding apparatus from the Late Jurassic of Germany, „PalZ”, 97 (2), 2023, s. 383–424, DOI: 10.1007/s12542-022-00644-4, ISSN 1867-6812 [dostęp 2024-01-08] (ang.).
- ↑ Andrea Villa, Loredana Macaluso, Thomas Mörs, Miocene and Pliocene amphibians from Hambach (Germany): new evidence for a late Neogene refuge in northwestern Europe, „Palaeontologia Electronica”, 27, Palaeontological Association, 2024, s. 1–56, DOI: 10.26879/1323 [dostęp 2024-01-08] (ang.).
- ↑ Dziennik Polska-Europa-Świat, 9.01.2009 r., s. 20.
- ↑ Xiaolin Wang, Zhonghe Zhou, Pterosaur embryo from the Early Cretaceous, „Nature”, 429 (6992), 2004, s. 621–621, DOI: 10.1038/429621a, ISSN 1476-4687 [dostęp 2024-01-06] (ang.).
- ↑ S. Christopher Bennett. New smallest specimen of the pterosaur Pteranodon and ontogenetic niches in pterosaurs. „Journal of Paleontology”. 92 (2), s. 254–271, 2018. DOI: 10.1017/jpa.2017.84. (ang.).
- ↑ Efficient flapping flight of pterosaurs - ProQuest [online], www.proquest.com [dostęp 2024-01-14] (pol.).
- kategoria
- wikiprojekt
| Geochronologia | |
|---|---|
| Wymarłe zwierzęta | |
| Rośliny kopalne | |
| Pojęcia |
