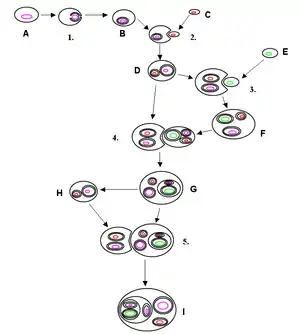
kolory oznaczają: czarny – błona komórkowa, różowy – eukariotyczny DNA, zielony – cyjanobakterie lub ctDNA, czerwony protobakterie lub mtDNA
Teoria endosymbiozy – teoria stanowiąca, że mitochondria, plastydy (jak chloroplasty) i być może inne organella komórki eukariotycznej powstały na skutek endosymbiozy pomiędzy różnymi mikroorganizmami. Zgodnie z nią niektóre organella pochodzą od wolno żyjących bakterii, które dostały się do innych komórek jako endosymbionty. Mitochondria rozwinęły się więc z proteobakterii (w szczególności zaś z Rickettsiales, kladu SAR11[1][2] lub ich bliskich krewnych), chloroplasty zaś od sinic.
Historia
Wyraz pochodzi z greki: ἔνδον (endon) oznacza „wewnątrz”, σύν (syn) – „razem”, βίωσις (biosis) zaś – „życie”. Teorię tę zaprezentował po raz pierwszy rosyjski botanik Konstantin Mierieżkowski w 1905[3][4]. Mierieżkowski zapoznał się z pracą innego botanika Andreasa Schimpera, który zaobserwował w 1883 podział chloroplastu w roślinie zielonej, podział znacznie przypominający ten spotykany u wolno żyjących sinic. Naukowiec ten zaproponował wstępnie w formie przypisu, że rośliny zielone powstały na skutek symbiozy dwóch organizmów[5].
W latach 20. XX wieku Ivan Wallin rozszerzył ten pomysł endosymbiotycznego pochodzenia na mitochondria[6]. Teorie te początkowo odrzucano bądź ignorowano. Dokładniejsze badania porównawcze pomiędzy sinicami i chloroplastami z użyciem mikroskopii elektronowej (jak choćby prace Hansa Risa[7]) łączyły się w całość z odkryciem, że plastydy i mitochondria posiadają swe własne DNA[8] (na tym etapie rozpoznane jako materiał dziedziczny organizmów), doprowadziły one do przywrócenia hipotezy w latach sześćdziesiątych.
Teoria endosymbiozy posunęła się do przodu i została poparta argumentami natury mikrobiologicznej w pracy Lynn Margulis z 1967, noszącej tytuł The Origin of Mitosing Eukaryotic Cells („Powstanie dzielących się mitotycznie komórek eukariotycznych”)[9]. W jej pracy z 1981 Symbiosis in Cell Evolution („Symbioza w ewolucji komórek”) argumentowała, że komórki eukariotyczne powstały ze zbiorowisk wpływających na siebie wzajemnie jednostek, wliczając w to endosymbiotyczne krętki, które rozwinęły się w eukariotyczne witki i rzęski. Ten ostatni pomysł nie zdobył sobie znacznej akceptacji, ponieważ wić nie posiada DNA ani nie wykazuje podobieństw ultrastrukturalnych z bakteriami czy archeonami. Zgodnie z pracą L. Margulis i D. Sagana[10] „życie nie przejęło globu poprzez walkę, ale przez tworzenie powiązań”, choćby przez współpracę. Rozważano także możliwość endosymbiotycznego pochodzenia peroksysomów, choć nie mają one DNA. Christian de Duve zaproponował, że mogą one być pierwszymi endosymbiontami, pozwalającymi komórkom wytrzymywać wzrastające ilości tlenu w ziemskiej atmosferze. Obecnie wydają się jednak tworzonymi de novo, niezgodnie z pomysłem ich symbiotycznego rodowodu. Choćby praca Gabaldón i współpracowników z 2006 podaje dowody przeciwko endosymbiotycznemu pochodzeniu peroksysomów, zamiast tego sugerując ich wyewoluowanie z siateczki śródplazmatycznej[11]. Kolejnym organellum o pochodzeniu endosymbiotycznym jest nitroplast stwierdzony u haptofita Braarudosphaera bigelowii. Pochodzi on od diazotroficznej sinicy, a początkowo układ ten uważano za zwykłą symbiozę[12].
Panuje pogląd, że przez tysiąclecia część DNA endosymbiontów została przetransportowana do jądra komórki ich gospodarza podczas ewolucyjnego przejścia od symbiotycznej wspólnoty organizmów do komórki eukariotycznej (seryjna endosymbioza).
Od symbiontów do organelli
Keeling i Archibald[13] opisują zwykłe kryterium różnicujące organella od endosymbiontów: zredukowany rozmiar genomu. W trakcie ewolucji endosymbiontów do organelli większość z ich genów przeniosła się do genomu gospodarza. Jego komórka i organellum musiały więc wykształcić mechanizm transportujący produkty białkowe potrzebne w organellum, jednakże produkowane teraz w innych częściach komórki gospodarza. Jednakże podnosząc przykład słodkowodnej ameby Paulinella chromatophora, zawierającej chromatofory uważane za powstałe z sinic, wymienieni autorzy stwierdzają, że nie jest to jedyne możliwe kryterium. Inne polega na przejęciu przez gospodarza kontroli nad regulacją podziału dawnego symbionta, na utrzymywaniu jego synchronizacji z własnym podziałem komórkowym[13]. Nowack i współpracownicy[14] zsekwencjonowali geny chromatoforu o długości 1,02 Mb i znaleźli tylko 867 białek kodowanych przez te fotosyntezujące komórki. Porównania z ich najbliższymi wolno żyjącymi krewnymi, sinicami z rodzaju Synechococcus o genomie długości 3 Mb zawierającym 3300 genów, ujawniły drastyczne skrócenie genomu chromatoforów. Te ostatnie dysponowały genami związanymi z fotosyntezą. Brakowało natomiast genów odpowiedzialnych za inne funkcje anaboliczne. Oznacza to znaczny stopień zależności wzrostu i przeżycia w ogóle endosymbiontów od swych gospodarzy. Chromatofory okazały się wobec tego niefunkcjonalne w obliczu celów specyficznych dla organelli w porównaniu z mitochondriami czy plastydami. Ta różnica mogła pobudzać wczesną ewolucję organelli fotosyntetycznych.
Dowody
Za powstaniem mitochondriów i plastydów z bakterii przemawiają następujące argumenty[15][16][17]:
- nowe mitochondria i plastydy tworzą się wyłącznie w procesie przypominającym podział
- w przypadku niektórych glonów, jak Euglena, plastydy można zniszczyć (np. użyciem pewnych środków chemicznych lub wydłużonym brakiem światła) bez wytworzenia jakichkolwiek innych szkód komórce. W takim wypadku plastydy nie zregenerują się. Świadczy to o tym, że ich regeneracja polegać musi na pozakomórkowym źródle, jak podział komórkowy czy endosymbioza
- organella te otacza podwójna błona biologiczna, przy czym jej błona wewnętrzna różni się składem od innych błon komórki eukariotycznej
- mitochondria i plastydy posiadają DNA różniące się od DNA jądrowego, przypominające natomiast DNA spotykane u bakterii, zarówno rozmiarami, jak i kolistym kształtem
- analiza sekwencji DNA i filogenetyka sugerują, że DNA jądrowe prawdopodobnie obejmuje również geny pochodzące z plastydów
- rybosomy tych organelli przypominają rybosomy bakteryjne (70S)
- w białkach pochodzących z tych organelli, jak i w proteinach bakteryjnych, początkowym aminokwasem jest N-formylometionina
- wiele cech struktury wewnętrznej i biochemii plastydów, jak na przykład obecność tylakoidów, a zwłaszcza chlorofilu, przypomina te spotykane u sinic. Badania filogenetyczne obejmujące genomy plastydów, bakterii i eukariotów również wskazują na bliskość plastydów i sinic
- mitochondria posiadają kilka enzymów i systemów transportu podobnych do bakteryjnych
- niektóre białka kodowane przez materiał genetyczny jądrowy są transportowane do organelli, a zarówno mitochondria, jak i plastydy posiadają jak na bakterie małe genomy. wiąże się to z wzrastającą zależnością od eukariotycznego gospodarza po wejściu w endosymbiozę. Większość genów genomów organelli została przez nie utracona albo przeniesiona do jądra. Większość genów potrzebnych mitochondriom czy plastydom znajduje się obecnie w jądrze komórkowym. Wiele z nich pochodzi właśnie z bakteryjnego endosymbionta
- plastydy spotyka się wśród bardzo różnych grup protistów, z których część wykazuje bliskie pokrewieństwo z formami nie dysponującymi plastydami. Sugeruje to, że jeśli chloroplasty powstały de novo, musiałyby to zrobić wiele razy, tak wiele, że trudno byłoby wyjaśnić bliskie podobieństwo jednych do drugich
- wiele z tych protistów zawiera plastydy „pierwotne”, których nie mogły uzyskać z innych eukariotów zawierających plastydy
- wśród eukariotów, które posiadły plastydy bezpośrednio od bakterii (znanych jako Archaeplastida), glaukocystofity mają chloroplasty silnie przypominające sinice. W szczególności pomiędzy ich dwoma błonami leży peptydoglikan
Endosymbioza wtórna
Pierwotna endosymbioza wiąże się z pochłonięciem bakterii przez inny organizm wolno żyjący. Endosymbioza wtórna zachodzi, gdy produkt endosymbiozy pierwotnej ulega wchłonięciu i zachowaniu przez inny wolno żyjący organizm eukariotyczny. Zjawisko to zachodziło kilka razy i doprowadziło do powstania bardzo zróżnicowanych grup glonów i innych eukariontów. Pewne organizmy doświadczają oportunistycznej korzyści z podobnego procesu, gdy, wchłonąwszy glon, czerpią z produktów jego fotosyntezy, jednakże gdy upolowana zdobycz umiera lub jest tracona, były gospodarz wraca do poprzedniego stylu życia. Obligatoryjne wtórne endosymbionty stają się zależne od swych organelli i nie potrafią przeżyć w sytuacji ich nieobecności[18]. RedToL, Red Algal Tree of Life Initiative stworzony przez National Science Foundation, podkreśla rolę krasnorostów w ewolucji na Ziemi odgrywaną poprzez wtórną endosymbiozę.
Przypadek tworzącej się wtórnej endosymbiozy mieli okazję obserwować Okamoto & Inouye (2005). Heterotroficzny protist Hatena arenicola zachowuje się w stosunku do zielenic jako drapieżnik do czasu, gdy je wchłonie. Następnie glon traci rzęski i cytoszkielet, a Hatena zostaje gospodarzem, przełączając się na zdobywanie pożywienia poprzez fotosyntezę i zdobywając umiejętność podążania za światłem. Traci też swój aparat pokarmowy.
Pomimo różnorodności organizmów zawierających plastydy, morfologia, biochemia, organizacja genomu i filogeneza molekularna plastydowego RNA i białek wskazuje na pojedyncze powstanie wszystkich obecnie istniejących plastydów, chociaż stanowi to nadal przedmiot dyskusji[19][20].
Niektóre gatunki, jak wesz ludzka, mają w mitochondriach liczne chromosomy. W połączeniu z filogenetyką genów kodowanych przez mitochondrialny DNA sugeruje to, że przodków tychże mitochondriów nie łączyło wspólne pochodzenie[21].
Przypisy
- ↑ Mitochondria Share an Ancestor With SAR11, a Globally Significant Marine Microbe. ScienceDaily, 2011-07-225. [dostęp 2011-07-26]. (ang.).
- ↑ J. Cameron Thrash et al.. Phylogenomic evidence for a common ancestor of mitochondria and the SAR11 clade. „Scientific Reports”. 1 (art. nr 13), 2011. DOI: 10.1038/srep00013. (ang.).
- ↑ Mereschkowski C. Über Natur und Ursprung der Chromatophoren im Pflanzenreiche. „Biol Centralbl”. 25 (18), s. 593–604, 1905. (niem.).
- ↑ Martin, William, Kowallik, Klaus V. Annotated English translation of Mereschkowsky’s 1905 paper „Über Natur und Ursprung der Chromatophoren im Pflanzenreiche”. „European Journal of Phycology”. 34 (03), s. 287–295, 1999. (ang.).
- ↑ Schimper AFW. Über die Entwicklung der Chlorophyllkörner und Farbkörper. „Bot. Zeitung”. 41 (7–10), s. 105–114, 121–131, 137–146, 153–162, 1883. (niem.).
- ↑ Wallin IE. The Mitochondria Problem. „The American Naturalist”. 57 (650), s. 255–261, 1923. DOI: 10.1086/279919. (ang.).
- ↑ Ris H, Singh RN. Electron microscope studies on blue-green algae. „J Biophys Biochem Cytol”. 9 (1), s. 63–80, 01 1961. DOI: 10.1083/jcb.9.1.63. PMID: 13741827. (ang.).
- ↑ Stocking C and Gifford E. Incorporation of thymidine into chloroplasts of Spirogyra. „Biochem. Biophys. Res. Comm.”. 1 (3), s. 159–164, 1959. DOI: 10.1016/0006-291X(59)90010-5. (ang.).
- ↑ Lynn Sagan. On the origin of mitosing cells. „J Theor Bio.”. 14 (3), s. 255–274, 1967. DOI: 10.1016/0022-5193(67)90079-3. PMID: 11541392. (ang.).
- ↑ Margulis, Lynn, Sagan, Dorion. Marvellous microbes. „Resurgence”. 206, s. 10–12, 2001. (ang.).
- ↑ Gabaldón T, Snel B, van Zimmeren F, Hemrika W, Tabak H, Huynen MA. Origin and evolution of the peroxisomal proteome. „Biol. Direct”. 1 (1), s. 8, 2006. DOI: 10.1186/1745-6150-1-8. PMID: 16556314. (ang.).
- ↑ Ramon Massana, The nitroplast: A nitrogen-fixing organelle, „Science”, 384 (6692), 2024, s. 160–161, DOI: 10.1126/science.ado8571, ISSN 0036-8075 [dostęp 2024-04-16] (ang.).
- 1 2 P.J. Keeling, Archibald, J.M. Organelle evolution: what’s in a name?. „Current Biology”. 18, s. 345–347, 2008. DOI: 10.1016/j.cub.2008.02.065. PMID: 18430636. (ang.).
- ↑ E.C. Nowack, Melkonian, M. & Glockner, G. Chromatophore genome sequence of Paulinella sheds light on acquisition of photosynthesis by eukaryotes. „Current Biology”. 18, s. 410–418, 2008. DOI: 10.1016/j.cub.2008.02.051. PMID: 18356055. (ang.).
- ↑ John W. Kimball: Kimball’s Biology Pages. 2010. [dostęp 2010-10-13]. [zarchiwizowane z tego adresu (2017-06-22)].
- ↑ Reece, J., Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson, 2010. Campbell Biology. 9th Edition Benjamin Cummings; 9th Ed. (2010-10-07).
- ↑ Raven, P., George Johnson, Kenneth Mason, Jonathan Losos, Susan Singer, 2010. Biology. McGraw-Hill 9th Ed. (2010-01-14).
- ↑ McFadden GI. Primary and secondary endosymbiosis and the origin of plastids. „J Phycology”. 37 (6), s. 951–959, 2001. DOI: 10.1046/j.1529-8817.20011126.x. (ang.).
- ↑ McFadden GI, van Dooren GG. Evolution: red algal genome affirms a common origin of all plastids. „Curr. Biol.”. 14 (13), s. R514–6, July 2004. DOI: 10.1016/j.cub.2004.06.041. PMID: 15242632. (ang.).
- ↑ Gould SB, Waller RF, McFadden GI. Plastid evolution. „Annu Rev Plant Biol”. 59 (1), s. 491–517, 2008. DOI: 10.1146/annurev.arplant.59.032607.092915. PMID: 18315522. (ang.).
- ↑ K. Georgiades, D. Raoult. The rhizome of Reclinomonas americana, Homo sapiens, Pediculus humanus and Saccharomyces cerevisiae mitochondria. „Biol Direct”. 6, s. 55, 2011. DOI: 10.1186/1745-6150-6-55. PMID: 22014084. PMCID: PMC3214132.
Bibliografia
- Alberts, Bruce: Molecular biology of the cell. New York: Garland Science, 2002. ISBN 0-8153-3218-1. (ang.).
- Blanchard JL, Lynch M. Organellar genes: why do they end up in the nucleus?. „Trends Genet.”. 16 (7), s. 315–320, July 2000. DOI: 10.1016/S0168-9525(00)02053-9. PMID: 10858662. (ang.).
- Jarvis P. Intracellular signalling: the chloroplast talks!. „Curr. Biol.”. 11 (8), s. R307–10, April 2001. DOI: 10.1016/S0960-9822(01)00171-3. PMID: 11369220. (ang.).
- Brinkman FS, Cherkasov A, et al.. Evidence that plant-like genes in Chlamydia species reflect an ancestral relationship between Chlamydiaceae, cyanobacteria, and the chloroplast. „Genome Res.”. 12 (8), s. 1159–1167, August 2002. DOI: 10.1101/gr.341802. PMID: 12176923. (ang.).
- Okamoto N, Inouye I. A secondary symbiosis in progress?. „Science”. 310 (5746), s. 287, October 2005. DOI: 10.1126/science.1116125. PMID: 16224014. (ang.).
- Cohen WD, Gardner RS: Viral Theory and Endosymbiosis. 1959. (ang.).