| Dromaeosauridae | |
| Matthew & Brown, 1922 | |
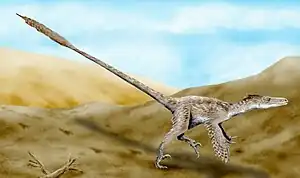 Velociraptor mongoliensis | |
| Systematyka | |
| Domena | |
|---|---|
| Królestwo | |
| Typ | |
| Podtyp | |
| Gromada | |
| Podgromada | |
| Infragromada | |
| Nadrząd | |
| Rząd | |
| Podrząd | |
| Infrarząd | |
| (bez rangi) | celurozaury |
| Nadrodzina | |
| Rodzina |
dromeozaury |
Dromeozaury (Dromaeosauridae) – rodzina teropodów należących do grupy Maniraptora. Często potocznie nazywane „raptorami”. Określenie to spopularyzował film Park Jurajski Stevena Spielberga. Nazwa Dromaeosauridae oznacza „biegające jaszczury” – od greckich słów dromeus/δρομευς („biegacz”) i sauros/σαυρος („jaszczur”).
Dromeozaury były niewielkimi, zwinnymi drapieżnikami, które rozwinęły się w okresie środkowej jury.Dowodem na to jest wykopanie zębów ich w Wielkiej Brytanii z tego okresu[1][2]. Największy rozkwit przeżywając w kredzie[3][4].
Ich rozmiary mieściły się w przedziale 0,5 do 7 metrów długości. Cechą charakterystyczną dromeozaurów był zakrzywiony pazur, zamocowany na wygiętym do tyłu, drugim palcu każdej z tylnych kończyn. Wyjątkiem wśród dromeozaurów jest Balaur, który miał taki pazur również na powiększonym pierwszym palcu stopy[5]. Pazur ten służył do polowania, zwierzę prawdopodobnie skakało na swoją ofiarę, a następnie zadawało serię pchnięć (nie jak wcześniej myślano – cięć rozszarpujących ciało ofiary), które przebijały się przez narządy wewnętrzne i żyły, doprowadzając do śmierci ofiary. Ogony dromeozaurów były usztywnione przez ścięgna, co pomagało utrzymać równowagę w czasie biegu i skoku. Microraptor gui miał nawet ogon zakończony niewielkim „sterem” z piór, co pomagało mu w zwinnym poruszaniu się. Obecność piór udowodniono również u m.in. welociraptora[6].
Ich życie społeczne pozostaje przedmiotem kontrowersji. Razem ze skamieniałościami roślinożernych tenontozaurów kilkakrotnie znajdowano zęby deinonychów i skamieniałości grup tych drapieżników, które według jednej z interpretacji miały zginąć, atakując większą od siebie ofiarę; odkrycia te przyczyniły się do popularyzacji poglądu, że przynajmniej niektóre gatunki dromeozaurów polowały w zorganizowanych watahach, co umożliwiało im atakowanie nawet kilkakrotnie większych zwierząt[7]. Część naukowców kwestionowała jednak zasadność takiego założenia, twierdząc, że deinonychy mogły równie dobrze skupiać się wokół znalezionej padliny tenontozaurów, a analiza znanych skamieniałości nie pozwala ani na przyjęcie, ani na odrzucenie żadnej z tych hipotez[8]. Roach & Brinkman (2007) wprost zakwestionowali tezę o dromeozaurach współpracujących w stadzie podczas łowów; ich zdaniem, odkryte skamieniałości tych zwierząt wskazują, że ich sposób polowania i żerowania był bardziej zbliżony do tego spotykanego u waranów z Komodo. W myśl tej hipotezy znalezione przy zabitych tenontozaurach skamieniałości deinonychów były w istocie szczątkami osobników, które zostały zabite i częściowo pożarte przez większe osobniki swojego gatunku[9].
Li i współpracownicy (2008) opisali skamieniałe tropy dromeozaurów odkryte w prowincji Szantung w Chinach. Ślady pochodzą z wczesnej kredy; należą do dwóch ichnorodzajów – Dromaeopodus (ślady zostawione przez duże zwierzę, porównywalne z achillobatorem) i mniejszego Velociraptorichnus. Autorzy publikacji opisują m.in. reprezentujące ichnorodzaj Dromaeopodus tropy sześciu dromeozaurów. Wszystkie zostały zostawione przez zwierzęta porównywalnej wielkości, idące w jednym kierunku; tropy różnych osobników nie pokrywały się, co więcej – zwierzęta idąc, pozostawały przez cały czas w równej odległości od siebie nawzajem. Zdaniem autorów publikacji, taki układ śladów wskazuje, że dromeozaury, które je zostawiły, nie przechodziły przez teren, na którym odkryto tropy, niezależnie i w pewnych odstępach czasu, ale w tym samym czasie, jako grupa; znalezisko to, jak twierdzą autorzy, dowodzi więc, że przynajmniej niektóre dromeozaury żyły stadnie. Nie dowodzi to automatycznie, że również wspólnie polowały (tropy z Chin nie wskazują, by zwierzęta w chwili zostawiania śladów polowały); autorzy publikacji twierdzą jednak, że takiej możliwości nie można też wykluczyć[10].
Klasyfikacja
- Rodzina Dromaeosauridae
- Atrociraptor
- Dromaeosauroides
- Luanchuanraptor
- Mahakala
- Pyroraptor
- ?Unquillosaurus
- Variraptor
- podrodzina Dromaeosaurinae
- podrodzina Microraptorinae
- podrodzina Unenlagiinae
- podrodzina Velociraptorinae
- podrodzina Halszkaraptorinae
- Halszkaraptor[13]
- Hulsanpes[14]
- Mahakala[15]
- Natovenator[16]
Filogeneza

| Dromaeosauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Przypisy
- ↑ S.J. Metcalf i inni, A new Bathonian (Middle Jurassic) microvertebrate site, within the Chipping Norton Limestone Formation at Hornsleasow Quarry, Gloucestershire, „Proceedings of the Geologists' Association”, 103 (4), 1992, s. 321–342, DOI: 10.1016/S0016-7878(08)80129-0, ISSN 0016-7878 [dostęp 2023-11-08].
- ↑ Simon Wills, Charlie J. Underwood, Paul M. Barrett, Machine learning confirms new records of maniraptoran theropods in Middle Jurassic UK microvertebrate faunas, Philip Mannion (red.), „Papers in Palaeontology”, 9 (2), 2023, DOI: 10.1002/spp2.1487, ISSN 2056-2799 [dostęp 2023-11-08] (ang.).
- ↑ Sarah N. Davis i inni, New records of Theropoda from a Late Cretaceous (Campanian-Maastrichtian) locality in the Magallanes-Austral Basin, Patagonia, and insights into end Cretaceous theropod diversity, „Journal of South American Earth Sciences”, 122, 2023, s. 104163, DOI: 10.1016/j.jsames.2022.104163, ISSN 0895-9811 [dostęp 2023-11-08].
- ↑ Steven E. Jasinski, Robert M. Sullivan, Peter Dodson, New Dromaeosaurid Dinosaur (Theropoda, Dromaeosauridae) from New Mexico and Biodiversity of Dromaeosaurids at the end of the Cretaceous, „Scientific Reports”, 10 (1), 2020, s. 5105, DOI: 10.1038/s41598-020-61480-7, ISSN 2045-2322 [dostęp 2023-11-08] (ang.).
- ↑ Zoltán Csiki, Mátyás Vremir, Stephen L. Brusatte, Mark A. Norell. An aberrant island-dwelling theropod dinosaur from the Late Cretaceous of Romania. „Proceedings of the National Academy of Sciences”, 2010. DOI: 10.1073/pnas.1006970107. (ang.).
- ↑ Alan H. Turner, Peter J. Makovicky, Mark A. Norell. Feather quill knobs in the dinosaur Velociraptor. „Science”. 317 (5845), s. 1721, 2007. DOI: 10.1126/science.1145076. (ang.).
- ↑ W. Desmond Maxwell, John H. Ostrom. Taphonomy and paleobiological implications of Tenontosaurus-Deinonychus associations. „Journal of Vertebrate Paleontology”. 15 (4), s. 707–712, 1995. (ang.).
- ↑ Kenneth Carpenter. Evidence of predatory behavior by carnivorous dinosaurs. „Gaia”. 15, s. 135–144, 1998. (ang.).
- ↑ Brian T. Roach, Daniel L. Brinkman. A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs. „Bulletin of the Peabody Museum of Natural History”. 48 (1), s. 103–138, 2007. (ang.).
- ↑ Rihui Li i inni, Behavioral and faunal implications of Early Cretaceous deinonychosaur trackways from China, „Naturwissenschaften”, 3, 95, 2008, s. 185–191, DOI: 10.1007/s00114-007-0310-7 (ang.).
- ↑ Alan H. Turner, Shaena Montanari, Mark A. Norell, A New Dromaeosaurid from the Late Cretaceous Khulsan Locality of Mongolia, „American Museum Novitates”, 2020 (3965), 2021, DOI: 10.1206/3965.1, ISSN 0003-0082 [dostęp 2024-01-08].
- ↑ Scott Hartman i inni, A new paravian dinosaur from the Late Jurassic of North America supports a late acquisition of avian flight, „PeerJ”, 7, 2019, e7247, DOI: 10.7717/peerj.7247, ISSN 2167-8359, PMID: 31333906, PMCID: PMC6626525 [dostęp 2024-01-13].
- ↑ Andrea Cau i inni, Synchrotron scanning reveals amphibious ecomorphology in a new clade of bird-like dinosaurs, „Nature”, 552 (7685), 2017, s. 395–399, DOI: 10.1038/nature24679, ISSN 1476-4687 [dostęp 2023-12-23] (ang.).
- ↑ Halszka Osmólska, Hulsanpes perlei n.g. n.sp. (Deinonychosauria, Saurischia, Dinosauria) from the Upper Cretaceous Barun Goyot Formation of Mongolia, „Neues Jahrbuch für Geologie und Paläontologie - Monatshefte”, 1982, s. 440–448, DOI: 10.1127/njgpm/1982/1982/440 [dostęp 2023-12-23] (ang.).
- ↑ Alan H. Turner i inni, A Basal Dromaeosaurid and Size Evolution Preceding Avian Flight, „Science”, 317 (5843), 2007, s. 1378–1381, DOI: 10.1126/science.1144066, ISSN 0036-8075 [dostęp 2023-12-23] (ang.).
- ↑ Sungjin Lee i inni, A non-avian dinosaur with a streamlined body exhibits potential adaptations for swimming, „Communications Biology”, 5 (1), 2022, s. 1–9, DOI: 10.1038/s42003-022-04119-9, ISSN 2399-3642 [dostęp 2023-12-23] (ang.).
- ↑ Alan H. Turner, Diego Pol, Julia A. Clarke, Gregory M. Erickson i inni. A basal dromaeosaurid and size evolution preceding avian flight. „Science”. 317 (5843), s. 1378–1381, 2007. DOI: 10.1126/science.1144066. (ang.).
- kategoria
- klasyfikacja
- lista
- wikiprojekt
| Infrarzędy dinozaurów | |
|---|---|
| Era dinozaurów | |
| Dyscypliny powiązane |
