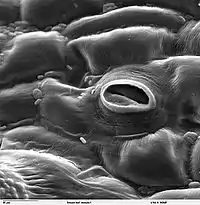


Aparat szparkowy i szparka (gr. stoma jama, szpara) – struktury wielokomórkowe (aparat) lub dwukomórkowe (szparka) pochodzenia epidermalnego roślin służące do kontrolowanej wymiany gazowej między tkankami wewnętrznymi rośliny i atmosferą[1].
Budowa
Aparaty szparkowe tworzą się we wczesnych stadiach rozwojowych rośliny, już na zawiązkach pędów. Składają się z dwóch komórek szparkowych, między którymi znajduje się otwór – szparka połączona do komory podszparkowej w miękiszu gąbczastym wewnątrz organu. Komórki szparkowe w odróżnieniu od pozostałych komórek epidermy zawierają chloroplasty i są zdolne do przeprowadzania fotosyntezy, nie posiadają natomiast plazmodesm. Na krawędziach komórek szparkowych (od strony szparki) wykształcają się występy ściany komórkowej zwane listwami szparkowymi. Gdy szparka jest zamknięta listwy zachodzą na siebie zamykając przepływ gazów. Komórki szparkowe wraz z listwami pokryte są grubą warstwą kutykuli. Ściany tych komórek są nierównomiernie zgrubiałe, zwłaszcza wzdłuż szparki.
U traw i części innych roślin jednoliściennych, zgrubienia te powodują, że komórki mają kształt hantli (w środku bywają tak mocno ściśnięte). Komórki szparkowe otoczone są kilkoma innymi komórkami epidermy nazywanymi komórkami przyszparkowymi. Biorą one udział w regulacji rozwarcia szparki.
Rośliny dwuliścienne, pozostałe jednoliścienne, nagonasienne oraz paprotniki i mchy nie posiadają komórek przyszparkowych, a cały aparat szparkowy składa się z dwóch komórek szparkowych. Najczęściej komórki te mają jednak kształt nerkowaty i tworzą owalny otwór. Zgrubienia ścian powodują, że zamknięta szparka jest szczelna, a mikrofibryle rozchodzące się promieniście od otworu umożliwiają rozciąganie komórki jedynie wzdłuż. W tym typie budowy aparatu szparkowego komórki szparkowe otoczone są bezpośrednio komórkami epidermy[2].
Aparat szparkowy może składać się z kilku komórek (aparaty szparkowe typu Gramineae, Amaryllidaceae i Mnium) lub kilkunastu (u wątrobowców). Aparaty te tworzone są przez komórki szparkowe i pomocnicze (przyszparkowe).
Rodzaje aparatów szparkowych[3]
- anomocytyczny – komórki szparkowe są otoczone zmienną liczbą komórek przyszparkowych, nie różniących się od komórek skórki
- anizocytyczny – komórki szparkowe są otoczone 3 komórkami przyszparkowymi, w tym jedną mniejszą
- diacytyczny – komórki szparkowe są otoczone 2 komórkami przyszparkowymi ułożonymi prostopadle do osi szparki
- paracytyczny – komórki szparkowe są otoczone 2 komórkami przyszparkowymi ułożonymi równolegle do osi szparki
- tetracytyczny – komórki szparkowe są otoczone 4 komórkami przyszparkowymi
Funkcja i mechanizm działania

Szparki mają decydujące znaczenie dla funkcjonowania układu wentylacyjnego roślin pozwalającego na dostarczanie niezbędnego dla procesu fotosyntezy dwutlenku węgla. Dzięki nim rośliny mogą skutecznie pobierać ten gaz chroniąc się równocześnie przed szkodliwymi stratami wody w wyniku transpiracji. Funkcjonowanie szparek wiąże się ze zdolnością do zmiany kształtu komórek szparkowych. Komórki te poprzez zmianę turgoru, wykorzystując zjawisko osmozy, powodują zamykanie (obniżenie turgoru) lub otwieranie (podwyższenie turgoru) szparek. Charakterystyczną cechą budowy komórek szparkowych jest nierównomierna grubość i specyficzna struktura ich ścian, związana z układem mikrofibryli celulozowych. W zwykłych komórkach cylindrycznych mikrofibryle ułożone są poprzecznie w stosunku do głównej osi komórki wzdłuż jej podłużnej osi. W nerkowatych komórkach szparkowych mikrofibryle celulozowe rozchodzą się promieniście od otworu szparkowego. W związku z tym obwód szparki zostaje wzmocniony, a sama komórka wygina się na zewnątrz w trakcie rozwierania. U traw środkowe części komórek szparkowych oddalają się wówczas, gdy wzrasta objętość ich bulwkowatych zakończeń, zdolnych do rozciągania się, co pozwala na rozszerzenie szczeliny między komórkami. Dawniej sądzono, że zmiany turgoru komórek szparkowych są skutkiem hydrolizy i syntezy skrobi w chloroplastach tych komórek. Obecnie wiadomo, że ruchy szparek powodowane są zmianą ciśnienia osmotycznego kationów potasu pobieranych przez komórki szparkowe przy jednoczesnym wydzielaniu protonów. Pobieranie jonów K+ zależne jest od stężenia dwutlenku węgla wewnątrz liścia (od intensywności fotosyntezy, czyli pośrednio od światła, zaopatrzenia w wodę, temperatury). Transport jonów potasu do wakuoli zachodzi w wyniku aktywacji pompy elektrogenicznej. Światło niebieskie, absorbowane przez karotenoid zeaksantynę, znajdującą się w tylakoidach gran chloroplastu, aktywuje H+-ATPazę w plazmolemie komórek szparkowych. Na skutek wzbudzenia zeaksantyny przez fotony światła niebieskiego zostaje zapoczątkowana kaskada zdarzeń, w wyniku których sygnał zostaje przekazany do cytoplazmy, gdzie aktywuje kinazę serynowo-treoninową. Kinaza fosforyluje C-końcowy fragment H+-ATPazy, aktywując enzym. Przyłączenie małego (32 kDa) białka regulatorowego, 14-3-3, do ufosforylowanego końca C H+-ATPazy umożliwia stabilizację enzymu. Defosforylacja enzymu prowadzi do dysocjacji 14-3-3. H+-ATPaza, wykorzystując ATP wytwarza gradient protonowy przez błonę, co umożliwia transport K+ przez specyficzne kanały potasowe i akumulację tego kationu w wakuoli. Do wakuoli pobierane są też jony chloru (na drodze symportu Cl-/H+), co pozwala zobojętnić ładunek kationu nagromadzonego w wakuoli.
Innym czynnikiem wpływającym na otwieranie aparatów szparkowych są jony jabłczanowe. Anion jabłczanowy powstaje przez hydrolizę skrobi, natomiast Cl- transportowany jest z komórek sąsiednich. Wzrastające stężenie jonów obniża potencjał wodny komórki szparkowej wywołując napływ wody z komórek o większym potencjale, napływająca woda zwiększa turgor, duży turgor jest przyczyną wyginania się komórek szparkowych i otwierania szparek[4].
Zamykanie się aparatów szparkowych następuje też pod wpływem innych związków chemicznych, m.in. kwasu abscysynowego, którego stężenie w soku roślin rośnie znacznie w warunkach suszy[5][6].
Występowanie
Aparaty szparkowe występują u wątrobowców, iglastych i traw, szparki u paprotników i większości roślin nasiennych[7]. Najliczniejsze są na młodych roślinach, na liściach i łodygach roślin zielnych. W mniejszym zagęszczeniu aparaty szparkowe występują w epidermie listków okwiatu i owoców. Mogą występować także na pędowych organach podziemnych (np. na kłączach). Na liściach, gdzie jest ich najwięcej, ich gęstość może dochodzić nawet do 1000 na 1 mm².
Aparaty szparkowe i szparki najczęściej zlokalizowane są na spodniej stronie liścia. Do wyjątków należą rośliny wodne o liściach pływających (np. grzybienie białe), których szparki znajdują się na górnej stronie liścia. U traw, których liście wznoszą się często pionowo, aparaty szparkowe rozmieszczone są równomiernie po obydwu stronach liścia. Rośliny wodne, zanurzone, nie wykształcają tego typu struktur w ogóle.
Szparki wykształcają się na powierzchni liścia (łodygi itd.), czasem też w zagłębieniach lub niewielkich wzniesieniach, a niekiedy także w dużych zagłębieniach zwanych kryptami szparkowymi.
Liczba szparek a stężenie dwutlenku węgla
Liczba szparek w liściach jest odwrotnie proporcjonalna do stężenia dwutlenku węgla w atmosferze. Ich gęstość (liczba na jednostkę powierzchni) zmniejsza się 0,6% przy zwiększeniu stężenia dwutlenku węgla o 1 ppmv[8]. W związku z tym gęstość szparek (ang. stomatal index, stomatal frequency) lub indeks porów może być używany do ocen zmian klimatu (patrz efekt cieplarniany).
Różne
- Przy otwartych szparkach transpiracja jest niewiele mniejsza niż parowanie z otwartego lustra wody.
- Szparki i aparaty szparkowe jako naturalne otwory w skórce chroniącej rośliny przed szkodliwym wpływem środowiska zewnętrznego stanowią wrota inwazji patogenów. Na przykład zarodniki grzybów z rzędu rdzowców Uredinales kiełkują na powierzchni blaszki liściowej, po dotarciu do szparki tworzą przycistkę wciskającą się do wnętrza liścia. Zarodniki pływkowe przemieszczają się w kierunku szparki stymulowane przez substancje chemiczne wydzielane przez roślinę. Po dotarciu do szparki zarodnik nieruchomieje i kiełkuje, a strzępki grzyba rozwijają się dalej wewnątrz blaszki liściowej.
- Związkiem o kluczowym znaczeniu dla funkcjonowania aparatów szparkowych jest tlenek azotu. Ten sam związek, który w przypadku leku Viagra rozluźnia mięśnie naczyń krwionośnych u ludzi wspomagając erekcję, u roślin powoduje zamykanie się szparek. Naukowcy sądzą, że spryskiwanie roślin substancją uwalniająca tlenek azotu może zwiększyć odporność roślin na niedostatek wody w środowisku[9].
Zobacz też
Przypisy
- ↑ Z. Hejnowicz: Anatomia i histogeneza roślin naczyniowych. Warszawa: Wydawnictwo Naukowe PWN, 1955. ISBN 83-01-13825-4.
- ↑ Jan Kopcewicz, Stanisław Lewak, Halina Gabryś: Fizjologia roślin. Warszawa: Wydawnictwo Naukowe PWN, 2002, s. 218-222. ISBN 83-01-13753-3.
- ↑ Bolesław Broda: Zarys botaniki farmaceutycznej. Warszawa: PZWL, 1975, s. 86-87.
- ↑ Lincoln Taiz, Eduardo Zeiger: Plant Physiology (Loosleaf). Wyd. 5. Sinauer Associates, Inc., 2010, s. 413-417. ISBN 978-0-87893-511-6.
- ↑ PE. Kriedemann i inni, Abscisic Acid and stomatal regulation, „Plant Physiology”, 49 (5), 1972, s. 842-847, DOI: 10.1104/pp.49.5.842, PMID: 16658058, PMCID: PMC366062.
- ↑ Rana Munns, Rod W. King, Abscisic Acid is not the only stomatal inhibitor in the transpiration stream of wheat plants, „Plant Physiology”, 88 (3), 1988, s. 703-708, DOI: 10.1104/pp.88.3.703, PMID: 16666371, PMCID: PMC1055648.
- ↑ Zygmunt Hejnowicz, Anatomia i histogeneza roślin naczyniowych : organy wegetatywne, Wyd. nowe (3 zm.), Warszawa: Wydaw. Naukowe PWN, 2002, ISBN 83-01-13825-4, OCLC 68674659 [dostęp 2021-06-08].
- ↑ F Wagner i inni, A natural experiment on plant acclimation: lifetime stomatal frequency response of an individual tree to annual atmospheric CO2 increase, „Proceedings of the National Academy of Sciences of the United States of America”, 93 (21), 1996, s. 11705-11708, DOI: 10.1073/pnas.93.21.11705, PMID: 11607712, PMCID: PMC38122.
- ↑ Viagra a więdnięcie (wiw.pl)