| Magnocellular cell | |
|---|---|
| Details | |
| System | Visual system |
| Location | Lateral geniculate nucleus of the thalamus |
| Identifiers | |
| NeuroLex ID | nifext_42 |
| Anatomical terms of neuroanatomy | |
Magnocellular cells, also called M-cells, are neurons located within the magnocellular layer of the lateral geniculate nucleus of the thalamus. The cells are part of the visual system. They are termed "magnocellular" since they are characterized by their relatively large size compared to parvocellular cells.
Structure
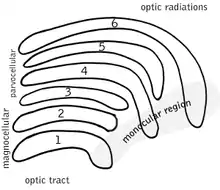
The full details of the flow of signaling from the eye to the visual cortex of the brain that result in the experience of vision are incompletely understood. Many aspects are subject to active controversy and the disruption of new evidence.[1][2]
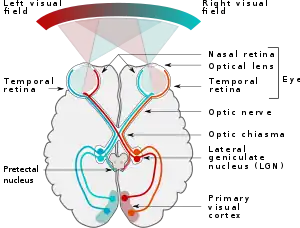
In the visual system, signals mostly travel from the retina to the lateral geniculate nucleus (LGN) and then to the visual cortex. In humans the LGN is normally described as having six distinctive layers. The inner two layers, (1 and 2) are magnocellular cell (M cell) layers, while the outer four layers, (3,4,5 and 6), are parvocellular cell (P cell) layers. An additional set of neurons, known as the koniocellular cell (K cell) layers, are found ventral to each of the M cell and P cell layers.[2][3]: 227ff [4] These layers were named this way because cells in the M layers of the LGN are larger than cells in the P layers.[3]: 228 [5]
M cells in the LGN receive input from parasol ganglion cells (which some neuroscientists call M cells),[3]: 226 and P cells receive input from midget retinal ganglion cells (which some neuroscientists call P cells).[3]: 226 [6][7]

From the LGN, the M pathway continues by sending information to the interblob regions of the 4Cα layer of the V1 region of the visual cortex, also called the "striate cortex".[6] Other cells in the striate are more influenced from signaling from P cells and yet others from K cells. As signals are passed to other regions of the cortex, the signals start to be less separate, more integrated, and more influenced by signals from other parts of the brain. While classically it is said that signaling through the M pathway ultimately flow out of the visual cortex through the dorsal stream and signaling through the P pathway ultimately flows to the ventral stream, subsequent studies have shown that both pathways influence both streams.[3]: 236
Function
The magnocellular pathway cannot provide finely detailed or colored information, but still provides useful static, depth, and motion information.[8][9] The M pathway has high light/dark contrast detection,[10] and is more sensitive at low spatial frequencies than high spatial frequencies. Due to this contrast information, M cells are essential for detecting changes in luminance, and performing visual search tasks and detecting edges.[11]
The M pathway is also important for providing information about the location of objects. M cells can detect the orientation and position of objects in space,[12] information that is sent through the dorsal stream.[13] This information is also useful for detecting the difference in positions of objects on the retina of each eye, an important tool in binocular depth perception.[14]
Cells in the M pathway have the ability to detect high temporal frequencies and can thus detect quick changes in the position of an object.[7] This is the basis for detecting motion.[10][15] The information sent to the intraparietal sulcus (IPS) of the posterior parietal cortex allows the M pathway to direct attention and guide saccadic eye movements to follow important moving objects in the visual field.[8][16][17] In addition to following objects with the eyes, the IPS sends information to parts of the frontal lobe that allows the hands and arms to adjust their movements to correctly grasp objects based on their size, position, and location.[13] This ability has led some neuroscientists to hypothesize that the purpose of the M pathway is not to detect spatial locations, but to guide actions related to the position and motion of objects.[18]
Some information has also been found to support the hypothesis that the M pathway is necessary for facial processing.[19]
Clinical significance
Abnormal magnocellular pathways and magnocellular cells can be associated with various disorders and ocular impairments, including dyslexia, prosopagnosia and schizophrenia.[8][15][19]
Dyslexia
Dyslexia is a disability which affects individual’s ability to read. It often first manifests in childhood, if at all; however, dyslexia can manifest itself in adulthood because of a brain tumor or lesion on/penetrating M cells.[15] There is no clear idea of the role of M cells and the magnocellular pathway in dyslexia.
One theory suggests that the nonlinearity, size, and compensation of miniature eye movements of M cells all help to focus on a single target and blur the surroundings, which is crucial in reading. This suggests that M cells are underdeveloped in many dyslexics. This may be due to genetics, autoimmunity, or nutrition. The KIAA0319 gene on chromosome six controls cell migration to the LGN during development; and studies in transgenic mice and on brains of people with dyslexia examined after they died, show malformations in the LGN and cells expressing KIAA0319 growing in the wrong place.[8] M cells are vulnerable to antineuronal antibodies which attack and render them unusable in the magnocellular pathway. This could be a cause of why dyslexics are more likely to have weakened immune systems.[8]
Another line of research suggests that defective eye movement caused by M cells is the cause of dyslexia. Since the magnocellular system is sensitive to image movement, and dyslexia is posited to be caused by abnormalities in M cells, dyslexics tend to focus on words longer, take shorter scans when reading, and stop more often per line. The study postulates that this is not caused by dyslexia but rather, low comprehension of the text causing abnormal eye movements in M cells. Therefore, it is difficult to conclude the importance of M cells in dyslexia from this study.[15]
Schizophrenia
Schizophrenia is a mental disorder in which people are unable to differentiate what is real and what is not. It is believed that the magnocellular pathway may help with facial recognition and discrimination in children, but when this pathway is not developed completely or correctly, facial processing is more difficult for individuals later in life. This is seen in people with schizophrenia and occurs when there are issues in the integration of information from the M cell and P cell pathways, making it difficult for individuals with schizophrenia to differentiate between reality and hallucinations.[19]
References
- ↑ Freud, E; Plaut, DC; Behrmann, M (October 2016). "'What' Is Happening in the Dorsal Visual Pathway". Trends in Cognitive Sciences. 20 (10): 773–84. doi:10.1016/j.tics.2016.08.003. PMID 27615805. S2CID 3053257.
- 1 2 Wallace, DJ; Fitzpatrick, D; Kerr, JN (25 January 2016). "Primate Thalamus: More Than Meets an Eye". Current Biology. 26 (2): R60–1. doi:10.1016/j.cub.2015.11.025. PMID 26811887.
- 1 2 3 4 5 Brodal, Per (2010). The central nervous system : structure and function (4th ed.). New York: Oxford University Press. ISBN 978-0-19-538115-3.
- ↑ Carlson, Neil R. (2007). Physiology of behavior (9th ed.). Boston: Pearson/Allyn & Bacon. ISBN 978-0205467242.
- ↑ NB: "Parvus means "small" in Latin, per Latin Dictionary and "magnus" means large, per Latin Dictionary
- 1 2 Callaway EM (July 2005). "Structure and function of parallel pathways in the primate early visual system". The Journal of Physiology. 566 (Pt 1): 13–9. doi:10.1113/jphysiol.2005.088047. PMC 1464718. PMID 15905213.
- 1 2 Nassi JJ, Callaway EM (May 2009). "Parallel processing strategies of the primate visual system". Nature Reviews. Neuroscience. 10 (5): 360–72. doi:10.1038/nrn2619. PMC 2771435. PMID 19352403.
- 1 2 3 4 5 Stein J (2014-01-01). "Dyslexia: the Role of Vision and Visual Attention". Current Developmental Disorders Reports. 1 (4): 267–280. doi:10.1007/s40474-014-0030-6. PMC 4203994. PMID 25346883.
- ↑ Jeffries, AM; Killian, NJ; Pezaris, JS (February 2014). "Mapping the primate lateral geniculate nucleus: a review of experiments and methods". Journal of Physiology, Paris. 108 (1): 3–10. doi:10.1016/j.jphysparis.2013.10.001. PMC 5446894. PMID 24270042.
- 1 2 Pokorny J (July 2011). "Review: steady and pulsed pedestals, the how and why of post-receptoral pathway separation". Journal of Vision. 11 (5): 7. doi:10.1167/11.5.7. PMID 21737512.
- ↑ Cheng A, Eysel UT, Vidyasagar TR (October 2004). "The role of the magnocellular pathway in serial deployment of visual attention". The European Journal of Neuroscience. 20 (8): 2188–92. doi:10.1111/j.1460-9568.2004.03675.x. PMID 15450098. S2CID 31456201.
- ↑ Skottun BC, Skoyles JR (January 2011). "On identifying magnocellular and parvocellular responses on the basis of contrast-response functions". Schizophrenia Bulletin. 37 (1): 23–6. doi:10.1093/schbul/sbq114. PMC 3004196. PMID 20929967.
- 1 2 Hebart MN, Hesselmann G (June 2012). "What visual information is processed in the human dorsal stream?". The Journal of Neuroscience. 32 (24): 8107–9. doi:10.1523/JNEUROSCI.1462-12.2012. PMC 6703654. PMID 22699890.
- ↑ Poggio GF, Poggio T (1984). "The analysis of stereopsis". Annual Review of Neuroscience. 7: 379–412. doi:10.1146/annurev.ne.07.030184.002115. PMID 6370081. S2CID 18838554.
- 1 2 3 4 Vidyasagar TR (January 2004). "Neural underpinnings of dyslexia as a disorder of visuo-spatial attention". Clinical & Experimental Optometry. 87 (1): 4–10. doi:10.1111/j.1444-0938.2004.tb03138.x. PMID 14720113. S2CID 45931933.
- ↑ Jayakumar J, Dreher B, Vidyasagar TR (May 2013). "Tracking blue cone signals in the primate brain" (PDF). Clinical & Experimental Optometry. 96 (3): 259–66. doi:10.1111/j.1444-0938.2012.00819.x. hdl:11343/39658. PMID 23186138. S2CID 32259651.
- ↑ Burr DC, Morrone MC, Ross J (October 1994). "Selective suppression of the magnocellular visual pathway during saccadic eye movements". Nature. 371 (6497): 511–3. Bibcode:1994Natur.371..511B. doi:10.1038/371511a0. PMID 7935763. S2CID 4332938.
- ↑ Goodale MA, Westwood DA (2004). "An evolving view of duplex vision: separate but interacting cortical pathways for perception and action". Current Opinion in Neurobiology. 14 (2): 203–11. doi:10.1016/j.conb.2004.03.002. PMID 15082326. S2CID 15917985.
- 1 2 3 Bortolon C, Capdevielle D, Raffard S (June 2015). "Face recognition in schizophrenia disorder: A comprehensive review of behavioral, neuroimaging and neurophysiological studies". Neuroscience and Biobehavioral Reviews. 53: 79–107. doi:10.1016/j.neubiorev.2015.03.006. PMID 25800172. S2CID 20887067.