| Gobiconodontidae Temporal range: | |
|---|---|
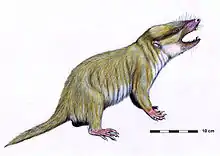 | |
| Life restoration of Gobiconodon | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Clade: | †Eutriconodonta (?) |
| Family: | †Gobiconodontidae Jenkins Jr. & Schaff, 1988 |
| Genera | |
| |
Gobiconodontidae is a family of extinct mammals that ranged from the mid-Jurassic to the early Late Cretaceous, though most common during the Early Cretaceous.[6] The Gobiconodontids form a diverse lineage of carnivorous non-therian mammals, and include some of the best preserved Mesozoic mammal specimens.
Biology
Like many other non-therian mammals, gobiocontids retained classical mammalian synapomorphies like epipubic bones (and likely the associated reproductive constrictions), venomous spurs and sprawling limbs. Through Spinolestes we also know that they had fur similar to that of modern mammals, with compound hair follicles with primary and secondary hairs. Spinolestes also possesses a clear diaphragm like modern mammals, as well as spines, dermal scutes and an ossified Meckel's cartilage. Furthermore, it may also display signs of dermatophytosis, suggesting that gobiconodontids, like modern mammals, were vulnerable to this type of fungal infection.[5]
Gobiconodontid dentition, being classically triconodont, has no analogue among living mammals, so comparisons are difficult. Like amphilestids but unlike triconodontids, occlusion happens by the molars essentially interlocking, with lower cusp "a" basically fitting between two upper molars.[6] Nonetheless, it's clear that most if not all gobiconodontids were primarily carnivorous, given the presence of long, sharp canines and incisors,[note 1] premolars with trenchant main cusps that were well suited to grasp and pierce prey, strong development of the mandibular abductor musculature, bone crushing ability in at least some species and several other features.[7]
Gobiconodontids are often among the largest mammals in Mesozoic faunal assemblages, with forms like Repenomamus and Gobiconodon exceeding 2 kilos. They were among the first mammals to be specialised to hunt vertebrate prey, and likely occupied the highest trophic levels among mammals in their faunal communities. Several forms like Gobiconodon and Repenomamus show evidence of scavenging, being among the few Mesozoic mammals to have significantly exploited that.[7] Evidence of predation on significantly larger dinosaurs is also known.[8]
At least in carnivorous niches, gobiconodontids were probably replaced by deltatheroidean metatherians, which are the dominant carnivorous mammals in Late Cretaceous faunal assemblages.[9] Competition between both groups is unattested, but in Asia the Early Cretaceous gobiconodontid diversity is replaced entirely by a deltatheroidean one, while in North America Nanocuris appears after the absence of Gobiconodon and other larger eutriconodonts.[10]
At least Spinolestes had xenarthrous vertebrae and osseous scutes, convergent to those of modern xenarthrans and to a lesser extent the hero shrew. This genus may have displayed an ecological role similar to that of modern anteaters, pangolins, echidnas, aardvark, aardwolf and numbat, being the second known Mesozoic mammal after Fruitafossor to have done so.[11]
Uniquely among crown-group mammals, gobiconodontids replaced their molariform teeth by successors of similar complexity, while in other mammals less complex replacements are the norm.[12]
Phylogeny
Cladogram after Marisol Montellano, James A. Hopson, James M. Clark (2008)[2] and Gao et al. (2010).[13]
| Gobiconodontidae |
| ||||||||||||||||||||||||
Notes
- ↑ In Gobiconodon, the canine-like lower incisors outrightly replace the true canines, which are vestigial.[7]
References
- ↑ Nao Kusuhashi; Yuan-Qing Wang; Chuan-Kui Li; Xun Jin (2019). "New gobiconodontid (Eutriconodonta, Mammalia) from the Lower Cretaceous Shahai and Fuxin formations, Liaoning, China". Vertebrata PalAsiatica. in press. doi:10.19615/j.cnki.1000-3118.190724.
- 1 2 Marisol Montellano; James A. Hopson; James M. Clark (2008). "Late Early Jurassic Mammaliaforms from Huizachal Canyon, Tamaulipas, México". Journal of Vertebrate Paleontology. 28 (4): 1130–1143. Bibcode:2008JVPal..28.1130M. doi:10.1671/0272-4634-28.4.1130. S2CID 128782275.
- 1 2 J.; Hu, Y.-M.; Wang, Y.-Q.; Li, C.-K. (2005). "A new triconodont (Mammalia) from the Early Cretaceous Yixian Formation of Liaoning, China". Vertebrata PalAsiatica. 43 (1): 1–10.
- ↑ J. Li; Y. Wang; Y. Wang; C. Li (2001). "A new family of primitive mammal from the Mesozoic of western Liaoning, China". Chinese Science Bulletin. 46 (9): 782–785. Bibcode:2001ChSBu..46..782L. doi:10.1007/BF03187223. ISSN 1001-6538. S2CID 129025369.
- 1 2 Martin, Thomas; Marugán-Lobón, Jesús; Vullo, Romain; Martín-Abad, Hugo; Luo, Zhe-Xi; Buscalioni, Angela D. (2015). "A Cretaceous eutriconodont and integument evolution in early mammals". Nature. 526 (7573): 380–384. Bibcode:2015Natur.526..380M. doi:10.1038/nature14905. PMID 26469049. S2CID 205245235.
- 1 2 Butler, P.M.; Sigogneau-Russell, D. (2016). "Diversity of triconodonts in the MiddleJurassic of Great Britain" (PDF). Palaeontologia Polonica. 67: 35–65. doi:10.4202/pp.2016.67_035 (inactive 1 August 2023).
{{cite journal}}: CS1 maint: DOI inactive as of August 2023 (link) - 1 2 3 Zofia Kielan-Jaworowska; Richard L. Cifelli; Zhe-Xi Luo (2004). "Chapter 7: Eutriconodontans". Mammals from the Age of Dinosaurs: origins, evolution, and structure. New York: Columbia University Press. pp. 216–248. ISBN 978-0-231-11918-4.
- ↑ Han, Gang; Mallon, Jordan C.; Lussier, Aaron J.; Wu, Xiao-Chun; Mitchell, Robert; Li, Ling-Ji (18 July 2023). "An extraordinary fossil captures the struggle for existence during the Mesozoic". Scientific Reports. 13 (1): 11221. Bibcode:2023NatSR..1311221H. doi:10.1038/s41598-023-37545-8. PMC 10354204. PMID 37464026.
- ↑ Zofia Kielan-Jaworowska; Richard L. Cifelli; Zhe-Xi Luo (2004). "Chapter 12: Metaherians". Mammals from the Age of Dinosaurs: origins, evolution, and structure. New York: Columbia University Press. pp. 425–462. ISBN 978-0-231-11918-4.
- ↑ Grossnickle, David M.; Polly, P. David (2013). "Mammal disparity decreases during the Cretaceous angiosperm radiation". Proceedings of the Royal Society B: Biological Sciences. 280 (1771). doi:10.1098/rspb.2013.2110. PMC 3790494. PMID 24089340. S2CID 6742591.
- ↑ Martin, Thomas; Marugán-Lobón, Jesús; Vullo, Romain; Martín-Abad, Hugo; Luo, Zhe-Xi; Buscalioni, Angela D. (2015). "A Cretaceous eutriconodont and integument evolution in early mammals". Nature. 526 (7573): 380–384. Bibcode:2015Natur.526..380M. doi:10.1038/nature14905. PMID 26469049. S2CID 205245235.
- ↑ Jäger, Kai R. K.; Cifelli, Richard L.; Martin, Thomas (2021). "Tooth eruption in the Early Cretaceous British mammal Triconodon and description of a new species". Papers in Palaeontology. 7 (2): 1065–1080. doi:10.1002/spp2.1329. S2CID 225501396.
- ↑ Chun-Ling Gao; Gregory P. Wilson; Zhe-Xi Luo; A. Murat Maga; Qingjin Meng & Xuri Wang (2010). "A new mammal skull from the Lower Cretaceous of China with implications for the evolution of obtuse-angled molars and 'amphilestid' eutriconodonts". Proceedings of the Royal Society B: Biological Sciences. 277 (1679): 237–246. doi:10.1098/rspb.2009.1014. PMC 2842676. PMID 19726475.
- Yuan, C.; Xu, L.; Zhang, X.; Xi, Y.; Wu, Y.; Ji, Q. (2009). "A new species of Gobiconodon (Mammalia) from western Liaoning, China and its implication for the dental formula of Gobiconodon". Acta Geologica Sinica. 83 (2): 207–211. doi:10.1111/j.1755-6724.2009.00035.x. S2CID 129667303.
- Thomas S. Kemp: The Origin & Evolution of Mammals. Oxford University Press, Oxford 2005, 331 Seiten, ISBN 0-19-850761-5.




