 | |
| Names | |
|---|---|
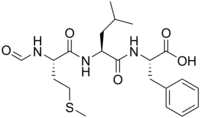
| Systematic IUPAC name
(2S)-2-{(2S)-2-[(2S)-2-Formamido-4-(methylsulfanyl)butanamido]-4-methylpentanamido}-3-phenylpropanoic acid | |
| Other names | |
| Identifiers | |
3D model (JSmol) |
|
| ChEBI | |
| ChemSpider | |
| MeSH | N-Formylmethionine+Leucyl-Phenylalanine |
PubChem CID |
|
| UNII | |
CompTox Dashboard (EPA) |
|
| |
| |
| Properties | |
| C21H31N3O5S | |
| Molar mass | 437.56 g·mol−1 |
Except where otherwise noted, data are given for materials in their standard state (at 25 °C [77 °F], 100 kPa).
Infobox references | |
N-Formylmethionyl-leucyl-phenylalanine (fMLF, fMLP or N-formyl-met-leu-phe) is an N-formylated tripeptide and sometimes simply referred to as chemotactic peptide is a potent polymorphonuclear leukocyte (PMN) chemotactic factor and is also a macrophage activator.[2]
fMLF is the prototypical representative of the N-formylated oligopeptide family of chemotactic factors. These oligopeptides are known to be, or mimic the actions of, the N-formyl oligopeptides that are (a) released by tissue bacteria, (b) attract and activate circulating blood leukocytes by binding to specific G protein coupled receptors on these cells, and (c) thereby direct the inflammatory response to sites of bacterial invasion. fMLF is involved in the innate immunity mechanism for host defense against pathogens.
fMLF led to the first discovery of a leukocyte receptor for a chemotactic factor, defined three different types of fMLF receptors that have complementary and/or opposing effects on inflammatory responses as well as many other activities, and helped define the stimulus-response coupling mechanisms by which diverse chemotactic factors and their G protein coupled receptors induce cellular function.
Discovery
In 1887, Élie Metchnikoff observed that leukocytes isolated from the blood of various animals were attracted towards certain bacteria.[3] This attraction was soon proposed to be due to soluble elements released by the bacteria [4] (see Harris[5] for a review of this area up to 1953). Peter Ward, Elmer Becker, Henry Showell, and colleagues showed that these elements were made by a variety of growing gram positive bacteria and gram negative bacteria and were of low molecular weight, i.e. below 3600 Dalton (unit)s.[6][7][8] Further studies by Schiffmann and colleges found that cultures of growing Escherichia coli released oligopeptides of between 150 and 1500 daltons that appeared to have a free Carboxylic acid group but not a free Amine group.[9]
Given these clues and knowledge that bacteria transcribe (see Transcription (genetics)) proteins starting with N-formylmethionine whereas eukaryotic cells mostly initiate protein synthesis with non-formylated methionine, Schiffmann, Corcoran, and Wahl theorized and then showed that N-formyl-methionine and a series N-formyl-methionyl dipeptides and tripeptides stimulated the chemotaxis of neutrophils isolated from rabbit peritoneal exudates as well as of macrophages isolated from guinea pig peritoneal exudates.[10] In further studies of various N-formylated oligopeptides, fMLF proved the most potent in stimulating rabbit neutrophil chemotaxis.[7] fMLF and a sampling of other, less potent, N-formyl oligopeptides were then found to stimulate a wide array of rabbit neutrophil functions such as: the transient auto-aggregation of these cells in suspension[11] and equally transient fall in circulating neutrophils when injected into rabbit veins (these responses result from an increase in neutrophil adhesiveness to each other and/or vascular endothelium);[12] the release (see degranulation) of intracellular granule-bound enzymes and other antimicrobial cytotoxic molecules; and the production and release of cytotoxic reactive oxygen species such as Superoxide and hydrogen peroxide.[7][13] All of these responses are part of the innate immune systems initial line of defense against bacterial invasions.
Follow-up studies found that the genes of mitochondria and chloroplasts organelles of Eukaryote cells, including those of humans, which, unlike nuclear genes, code for N-formyl-methionine proteins, release N-formyl-methionyl containing peptides with chemotactic activities that exactly mimic those of fMLF chemotaxis[14] These organelle-derived formylated peptides are true analogs of fMLF that operate through fMLF receptors to recruit circulating blood leukocytes to and thereby initiate inflammation responses at sites of cell damage and tissue destruction not caused by bacteria.[15] Thus, fMLF can act as a find-me signal, released by dead or dying cells to attract phagocytes to those cells, so that the phagocytes phagocytose the dead or dying cells, thereby clearing up the damage.[14] fMLF and other N-formylated oligopeptides were found to be similarly active in human neutrophils.[16][17] The high degree of structural specificity of a broad series of formylated peptides in stimulating these neutrophil responses, the specific binding of N-formylated oligopeptides to neutrophils with affinities that paralleled their stimulating potencies,[18] the ability of t-carbobenzoxy-phenylalanyl-methionine to bind to but not stimulate neutrophils and thereby to block the neutrophil binding and stimulating activity of N-formylated oligopeptides,[19][20] and the ability of the formylated oligopeptides to desensitize (i.e. render unresponsive) neutrophil functional responses to themselves but have no or a lesser ability to desensitize to a range of other chemotactic stimuli[21][22] provided strong suggestive evidence that the formylated peptides acted on cells through a common, dedicated receptor system that differed from other chemotactic factor receptors.
Receptors
The studies cited above lead to the eventual cloning of the human Formyl peptide receptor 1, a G protein coupled receptor that binds fMLF and other formylated oligopeptides to mediate their stimulatory actions on human and rabbit neutrophils. Subsequently, Formyl peptide receptor 2 and Formyl peptide receptor 3 were also cloned based on the similarities in their amino acid sequence to that of formyl peptide receptor 1. Formyl peptide receptors 2 and 3 have very different abilities to bind and respond to formylated oligopeptides including fMLF compared to formyl peptide receptor 1 and compared to each other and have very different functions than those of formyl peptide receptor 1.[23]
References
- ↑ n-formylmethionine leucyl-phenylalanine, Cancerweb
- ↑ Panaro MA, Mitolo V (Aug 1999). "Cellular responses to fMLF challenging: a mini-review". Immunopharmacology and Immunotoxicology. 21 (3): 397–419. doi:10.3109/08923979909007117. PMID 10466071.
- ↑ Metchnikoff E (1887). "Sur la lutte des cellules de l'organisme contre l'invasion des microbes". Ann. Inst. Pasteur. 1: 321.
- ↑ Grawitz P (1887). "unknown". Virchows Adz. IIO. I.
{{cite journal}}: Cite uses generic title (help) - ↑ Harris H (Jul 1954). "Role of chemotaxis in inflammation". Physiological Reviews. 34 (3): 529–62. doi:10.1152/physrev.1954.34.3.529. PMID 13185754.
- ↑ Ward PA, Lepow IH, Newman LJ (Apr 1968). "Bacterial factors chemotactic for polymorphonuclear leukocytes". The American Journal of Pathology. 52 (4): 725–36. PMC 2013377. PMID 4384494.
- 1 2 3 Showell HJ, Freer RJ, Zigmond SH, Schiffmann E, Aswanikumar S, Corcoran B, Becker EL (May 1976). "The structure-activity relations of synthetic peptides as chemotactic factors and inducers of lysosomal secretion for neutrophils". The Journal of Experimental Medicine. 143 (5): 1154–69. doi:10.1084/jem.143.5.1154. PMC 2190180. PMID 1262785.
- ↑ Becker EL, Showell HJ (Jun 1974). "The ability of chemotactic factors to induce lysosomal enzyme release. II. The mechanism of release". Journal of Immunology. 112 (6): 2055–62. doi:10.4049/jimmunol.112.6.2055. PMID 4825785.
- ↑ Schiffmann E, Showell HV, Corcoran BA, Ward PA, Smith E, Becker EL (Jun 1975). "The isolation and partial characterization of neutrophil chemotactic factors from Escherichia coli". Journal of Immunology. 114 (6): 1831–7. doi:10.4049/jimmunol.114.6.1831. PMID 165239. S2CID 22663271.
- ↑ Schiffmann E, Corcoran BA, Wahl SM (Mar 1975). "N-formylmethionyl peptides as chemoattractants for leucocytes". Proceedings of the National Academy of Sciences of the United States of America. 72 (3): 1059–62. Bibcode:1975PNAS...72.1059S. doi:10.1073/pnas.72.3.1059. PMC 432465. PMID 1093163.
- ↑ O'Flaherty JT, Kreutzer DL, Ward PA (Jul 1977). "Neutrophil aggregation and swelling induced by chemotactic agents". Journal of Immunology. 119 (1): 232–9. doi:10.4049/jimmunol.119.1.232. PMID 874320. S2CID 36465249.
- ↑ O'Flaherty JT, Showell HJ, Ward PA (May 1977). "Neutropenia induced by systemic infusion of chemotactic factors". Journal of Immunology. 118 (5): 1586–9. doi:10.4049/jimmunol.118.5.1586. PMID 858915. S2CID 32354288.
- ↑ Becker EL, Sigman M, Oliver JM (Apr 1979). "Superoxide production induced in rabbit polymorphonuclear leukocytes by synthetic chemotactic peptides and A23187". The American Journal of Pathology. 95 (1): 81–97. PMC 2042294. PMID 219701.
- 1 2 Carp H (Jan 1982). "Mitochondrial N-formylmethionyl proteins as chemoattractants for neutrophils". The Journal of Experimental Medicine. 155 (1): 264–75. doi:10.1084/jem.155.1.264. PMC 2186576. PMID 6274994.
- ↑ Dorward DA, Lucas CD, Chapman GB, Haslett C, Dhaliwal K, Rossi AG (May 2015). "The role of formylated peptides and formyl peptide receptor 1 in governing neutrophil function during acute inflammation". The American Journal of Pathology. 185 (5): 1172–84. doi:10.1016/j.ajpath.2015.01.020. PMC 4419282. PMID 25791526.
- ↑ O'Flaherty JT, Kreutzer DL, Ward PA (Mar 1978). "Chemotactic factor influences on the aggregation, swelling, and foreign surface adhesiveness of human leukocytes". The American Journal of Pathology. 90 (3): 537–50. PMC 2018255. PMID 564610.
- ↑ Lehmeyer JE, Snyderman R, Johnston RB (Jul 1979). "Stimulation of neutrophil oxidative metabolism by chemotactic peptides: influence of calcium ion concentration and cytochalasin B and comparison with stimulation by phorbol myristate acetate". Blood. 54 (1): 35–45. doi:10.1182/blood.V54.1.35.35. PMID 444673.
- ↑ Aswanikumar S, Corcoran B, Schiffmann E, Day AR, Freer RJ, Showell HJ, Becker EL (Jan 1977). "Demonstration of a receptor on rabbit neutrophils for chemotactic peptides". Biochemical and Biophysical Research Communications. 74 (2): 810–7. doi:10.1016/0006-291x(77)90375-8. PMID 836328.
- ↑ E. Schiffmann; B. A. Corcoran; A. Aswanikumar. (1978). "Molecular events in the response of neutrophils to synthetic N-formylmethionine chemotactic peptides.". In J. A. Gallin; P. G. Quie (eds.). Leukocyte chemotaxis: Methodology, physiology, clinical implications. New York.: Raven Press.
- ↑ O'Flaherty JT, Showell HJ, Kreutzer DL, Ward PA, Becker EL (Apr 1978). "Inhibition of in vivo and in vitro neutrophil responses to chemotactic factors by a competitive antagonist". Journal of Immunology. 120 (4): 1326–32. doi:10.4049/jimmunol.120.4.1326. PMID 641351. S2CID 42477532.
- ↑ O'Flaherty JT, Kreutzer DL, Showell HS, Becker EL, Ward PA (Dec 1978). "Desensitization of the neutrophil aggregation response to chemotactic factors". The American Journal of Pathology. 93 (3): 693–706. PMC 2018345. PMID 717543.
- ↑ O'Flaherty JT, Kreutzer DL, Showell HJ, Vitkauskas G, Becker EL, Ward PA (Mar 1979). "Selective neutrophil desensitization to chemotactic factors". The Journal of Cell Biology. 80 (3): 564–72. doi:10.1083/jcb.80.3.564. PMC 2110355. PMID 457760.
- ↑ Li Y, Ye D (Jul 2013). "Molecular biology for formyl peptide receptors in human diseases". Journal of Molecular Medicine. 91 (7): 781–9. doi:10.1007/s00109-013-1005-5. PMID 23404331. S2CID 150459.
External links
- N-Formylmethionine Leucyl-Phenylalanine at the U.S. National Library of Medicine Medical Subject Headings (MeSH)